 Reproduced, with permission, from:
De Fabo, E. C., and F. P. Noonan. 1986. Urocanic acid: On its role in the regulation of UVB-induced systemic immune suppression. In Stratospheric ozone. Vol. 2 of Effects of changes in stratospheric ozone and global climate, ed. J. G. Titus, 113-18. Proceedings of the United Nations Environment Programme (UNEP)/Environmental Protection Agency (EPA) International Conference on Health and Environmental Effects of Ozone Modification and Climate Change. Washington, D.C.: U.S. Environmental Protection Agency.
Reproduced, with permission, from:
De Fabo, E. C., and F. P. Noonan. 1986. Urocanic acid: On its role in the regulation of UVB-induced systemic immune suppression. In Stratospheric ozone. Vol. 2 of Effects of changes in stratospheric ozone and global climate, ed. J. G. Titus, 113-18. Proceedings of the United Nations Environment Programme (UNEP)/Environmental Protection Agency (EPA) International Conference on Health and Environmental Effects of Ozone Modification and Climate Change. Washington, D.C.: U.S. Environmental Protection Agency.
Edited by
James G. Titus
U.S. Environmental Protection Agency
The George Washington University Medical Center
Washington, DC USA
We have previously postulated that UVB-induced systemic immune suppression is mediated by a specific skin photoreceptor situated in the stratum corneum of mammalian skin. This photoreceptor is considered to be a biological signal transducer capable of converting UV-B radiation (290-320 nm) directly into a biochemical signal that then "switches-on" formation of antigen-specific suppressor cells by way of an "alteration" in antigen presentation.
The UVB-absorbing photoreceptor is tentatively identified as transurocanic acid (trans-UCA). The mode of immune regulation associated with it is suggested to be initiated by photoconversion to its cis isomer (cis UCA). Recent evidence is presented to support this concept showing that cis UCA in contrast to trans-UCA can induce an antigen-presenting "alteration" in splenic dendritic cells in the absence of any UV radiation.
A model is presented outlining a mechanism in which UCA-initiated immune suppression is involved with the prevention of autoimmune attack against "photoantigens" associated with sun-damaged skin. We propose that sunlight-induced skin tumor outgrowth may be an inadvertent consequence of this normally protective mechanism.
Urocanic acid (UCA, de-aminated histidine) is a major UVB-absorbing compound located in the stratum corneum. The naturally occurring form is the trans isomer; it isomerizes to the cis form on absorption of UV-B radiation.
Although the existence of UCA has been known for more than 100 years (Jaffe 1974), no physiological function for it has ever been established. Furthermore, even though UCA can attenuate ultraviolet (UV) radiation striking the skin of mammals, in effect acting as a photoprotector of DNA or as a sunscreen protecting against erythema, it may be playing a more fundamental physiological role as an immune-regulating skin photoreceptor (De Fabo and Noonan 1983). We discuss the evidence supporting this idea in this paper.
The ability of UV radiation to suppress a variety of immune responses in animals and humans has been well documented and reviewed (Kripke and Fisher 1976; Fisher and Kripke 1977; Morrison 1984; Parrish, Kripke, and Morrison 1984; Daynes and Krueger 1983). It has been shown that UV-B (290-320 nm) is responsible for suppressing the immunologic rejection response to transplanted UV-induced tumor (De Fabo and Kripke 1979, 1980).
Another major UV-B effect on immunity is its ability to suppress the delayed-type hypersensitivity (DTH) response to skin sensitizing antigens such as trinitrochlorobenzene (TNCB) in mice (Noonan, De Fabo, and Kripke, 1981a, 1981b). In this reaction mice that have been pretreated with UV radiation show a systemic suppression of the DTH response. We previously showed that it was energy primarily in the UV-B band that can diminish the DTH response in dose-dependent fashion (Noonan, De Fabo, and Kripke 1981a). The kinetics of this response was similar to the UVB-induced suppression of tumor rejection (De Fabo and Kripke 1979, 1980).
In further studies on the systemic suppression of DTH which were designed to elucidate the first light-absorbing event a relative wavelength effectiveness or "action" spectrum of this effect by UV radiation was estimated. The significant finding of this study was that radiation of wavelengths between 320 nm and 250 nm (UV-B and UV-C) exhibited differing effectiveness in suppressing the DTH response. This clear-cut differential in wavelength effectiveness suggested that a specific molecule, i.e., a "UVB-transducing" photoreceptor was absorbing this radiation and using it to "turn on" an immune suppressor response to the skin sensitizing chemical antigens. We suggested this mechanism might exist to "down regulate" the rejection response against new "photoantigens" put on skin cells by exposure to the UV-B component of sunlight. In effect this suppression would prevent uncontrolled autoimmune attack against sun-damaged cells (De Fabo and Noonan 1983). We found the shape of the action spectrum closely matched the absorption spectrum of UCA (De Fabo and Noonan 1983). Because of the close match between our in vivo derived action spectrum and the in vitro absorption spectrum of urocanic acid, we suggested that trans-UCA may be acting as the UVB-absorbing photoreceptor initiating events leading to immune suppression of DTH (De Fabo and Noonan 1983). Furthermore, because of close photobiological and immunological similarities to the suppression of tumor rejection, we suggested that perhaps this suppressive response to transplanted UV-induced skin tumors might also involve UCA (De Fabo and Noonan 1983).
One of the major puzzles about skin cancer is that nearly all UV-induced skin tumors in mice are highly antigenic and capable of evoking strong immunologic rejection responses by the host. Thus, it is unclear, given the strong immune response against these tumors, how they are able to grow out in the host.
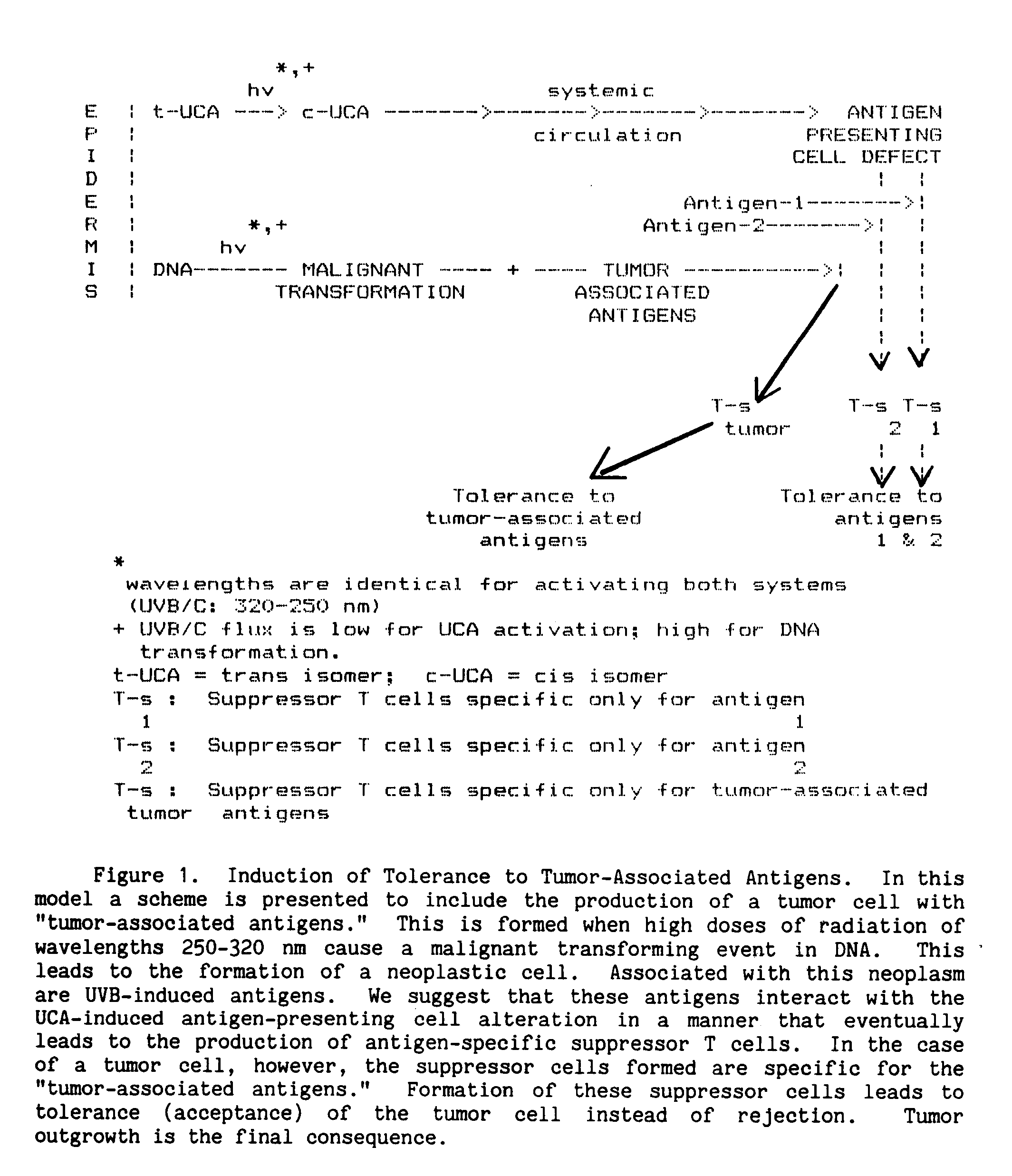
One possible explanation may involve an interaction between immune suppression mediated by UCA and tumor formation mediated by UV effects on DNA (De Fabo and Noonan 1984). Figure 1 presents a scheme for this type of interaction.
As previously noted, the underlying immunologic problem in mice irradiated with UV-B appears to be an alteration or "defect" in antigen presentation (Greene et al. 1979; Noonan et al. 1981). It seems that, because of this "defect", antigen-specific suppressor cells, rather than effector cells, are formed when antigen is given to a UVB-irradiated mouse. The hypothesis we put forward is that UCA may interact with antigen-presenting cells, e.g., macrophages, skin Langerhans cells, or splenic dendritic cells in such a way as to make defective their presentation of antigen to stimulator T cells. This process we propose is initiated by photoisomerization of trans-UCA to its highly soluble cis form. Once formed, the cis isomer or a secondary intermediate would interact with antigen-presenting cells in such a way as to alter their capacity to present antigen. This "defect" in antigen presentation (or processing) would become the signal for stimulation of suppressor cells rather than effector cells. The specificity of the suppressor cells would be determined by the type of antigen presented to the "defective" antigen-presenting cell (De Fabo and Noonan 1983).
This hypothesis has received strong support from the observations that (a) injection of cis UCA into mice induces an antigen-processing alteration in splenic dendritic cells without UV radiation; injection of trans-UCA was without effect (Noonan, De Fabo, and Morrison 1986) and (b) mice genetically deficient in histidase and thus in epidermal UCA showed no suppression of the CHS response when exposed to known immunosuppressive doses of UV-B (De Fabo et al. 1983).
In conclusion, it appears that a mechanism exists in skin that can directly bring about interaction between solar UV-B and the immune system in a way previously unknown. Further investigations into such a mechanism may open up new directions for determining the causes of skin cancer and other UVB immune-associated diseases.
For many years the physiological role for UCA has remained obscure, although a relatively large amount of this substance, in the trans configuration, accumulates in mammalian epidermis. UCA has been suggested to act in the skin as a natural sunscreen. Alternatively, we have proposed a physiological role for trans-UCA as a UVB-absorbing skin photoreceptor necessary to regulate against autoimmune attack on sun-damaged skin. In this capacity, trans-UCA is converted to the cis isomer that can then initiate the production of antigen-specific suppressor cells via an alteration in antigen-presentation. Thus, during prolonged sun exposure, suppressor cells form, which are specific for "photoantigens" induced on sun-damaged skin. These suppressor cells can prevent effector cells from carrying out an autoimmune attack against these "photoantigens."
Because such a mechanism exists, UCA may be involved in the outgrowth of sun-induced skin tumors, inadvertently protecting tumor cells from attack by effector cells. That is, due to the production of these "photoantigen"-specific suppressor cells that also recognize UV-induced antigens on tumors, tumor growth continues and the concentration of these UV-induced antigens steadily increases, keeping primed the production of specific suppressor cells (Figure 1). Without reversing this process, immune surveillance of the tumor (attack and destruction) will be prevented and tumor outgrowth will be the final outcome.
Daynes, R.A., and G. Krueger, eds. 1983. Experimental and clinical photoimmunology. Vol II. Cleveland, OH: CRC Press, Inc.
De Fabo, E.C., and M.L. Kripke. 1979. Dose-response characteristics of immunologic unresponsiveness to UV-induced tumors produced by UV irradiation of mice. Photochem. Photobiol. 30:385-390.
De Fabo, E.C., and M.L. Kripke. 1980. Wavelength dependence and dose-rate independence of UV radiation-induced suppression of immunologic unresponsiveness of mice to a UV-induced fibrosarcoma. Photochem. Photobiol. 32:183-188.
De Fabo, E.C., and F.P. Noonan. 1983. Mechanism of immune suppression by ultraviolet radiation in vivo. I: Evidence for the existence of a unique photoreceptor in skin and its role in photoimmunology. J. Exp. Med. 158:84-98.
De Fabo, E.C., and F.P. Noonan. 1984. Two-photoreceptor model for photocarcinogenesis. In Photobiology, 1984. Proceedings of the Ninth International Congress on Photobiology, eds. J. Jagger, J. Longworth, and W. Shropshire, J., 144. New York: Praeger Scientific.
De Fabo, E.C., F.P. Noonan, M. Fisher, J. Burns, and H. Kacser. 1983. Further evidence that the photoreceptor mediating UV-induced systemic immune suppression is urocanic acid. J. Invest. Dermatol. 80:319.
Fisher, M.S., and M.L. Kripke. 1977. Systemic alteration induced in mice by ultraviolet light irradiation and its relationship to ultraviolet carcinogenesis. Proc. Natl. Acad. Science USA. 74:1688-1692.
Greene, M.I., M.S. Sy, M.L. Kripke, and B. Benacerraf. 1979. Impairment of antigen-presenting cell function by ultraviolet radiation. Proc. Natl. Acad. Sci. USA. 76:6592-6595.
Jaffe, M. 1974. Concerning a new constituent of dog urine. Ber. Deut. Chem. Ges. 7:1669-1673.
Kripke, M.L., and M. Fisher. 1976. Immunologic parameters of ultraviolet carcinogenesis. J. Natl. Cancer Inst. 57:211-215.
Morrison, W.L. 1984. Photoimmunology. Photochem. Photobiol. 40:781-787.
Noonan, F.P., E.C. De Fabo, and M.L. Kripke. 1981a. Suppression of contact hypersensitivity by UV radiation and its relationship to UV-induced suppression of tumor immunity. Photochem. Photobiol. 34:683-690.
Noonan, F.P., E.C. De Fabo, and M.L. Kripke. 1981b. Suppression of contact hypersensitivity by ultraviolet radiation: An experimental model. Springer Seminars in Immunopathology. 4:293-304.
Noonan, F.P., E.C. De Fabo, and H. Morrison. 1986. Cis-urocanic acid, a UVB irradiation product, initiates an antigen presenting cell defect in vivo. Photochem. Photobiol. 43 supp:18s.
Noonan, F.P., M.L. Kripke, G.M. Pedersen, and M.I. Greene. 1981. Suppression of contact hypersensitivity in mice by ultraviolet irradiation is associated with defective antigen-presentation. Immunology. 43:527-533.
Parrish, J.A., M.L. Kripke, and W.L. Morrison, eds. 1984. Photoimmunology. New York: Plenum Medical Book Company.
{kind=link}