 Reproduced, with permission, from:
World Health Organization (WHO). 1989. Geographical distribution of arthropod-borne diseases and their principal vectors. Unpublished document WHO/VBC/89.967. Geneva: World Health Organization.
Reproduced, with permission, from:
World Health Organization (WHO). 1989. Geographical distribution of arthropod-borne diseases and their principal vectors. Unpublished document WHO/VBC/89.967. Geneva: World Health Organization.
Vector Biology and Control Division
CONTENTS
INTRODUCTION 1. Malaria G. B. White 2. Lymphatic filariasis G. B. White 3. Mosquito-borne virus diseases 3.1 Yellow fever M. G. R. Varma 3.2 Dengue and dengue haemorrhagic fever M. G. R. Varma 3.3 Japanese encephalitis M. G. R. Varma 3.4 Other encephalitides G.B. White 3.4.1 Californiaencephalitis 3.4.2 Eastern equine encephalitis 3.4.3 St Louis encephalitis 3.4.4 Western equine encephalitis 3.4.5 Venezuelan equine encephalitis 4. Tick-borne diseases M. G. R. Varma 4.1 Tick-borne virus diseases 4.1 .1 Colorado tick fever 4.1.2 Louping ill 4.1.3 Encephalitis 4.1.4 Kyasanur Forest disease 4.2 Tick-borne borrelioses 4.2.1 Tick-borne relapsing fever 4.2.2 Erythema borreliosis migrans 4.3 Tick-borne rickettsioses 4.3.1 Rocky Mountain spotted fever 4.3.2 Boutonneuse fever, Siberian and Queensland tick typhus 5. Mite-borne rickettsiosis M. G. R. Varma 6. Louse-borne diseases J. Maunder 7. The trypanosomiases 7.1 African D. M. Minter 7.2 American C. J. Schofield 8. The leishmaniases D. M. Minter 9. Flea-borne diseases I.A.H. Ismail/J. Maunder 9.1 Plague 9.2 Murine typhus 10. Onchocerciasis Anonymous 11. Loiasis Anonymous ACKNOWLEDGEMENTSINTRODUCTION
This manual provides information on the geographical distribution of the main diseases transmitted by mosquitos, ticks, mites, lice, tsetse flies, kissing bugs, phlebotomine sandflies, fleas, blackflies and deer flies. In addition, the distribution of the most important vectors is described in short texts, illustrated by over 100 maps*. Although much of this information can be obtained from various specialized reviews on each of the individual diseases or from comprehensive textbooks on tropical diseases, it is not easily available in a form suitable for the briefing of epidemiologists, health planners and medical entomologists, or for trainers and teachers involved in advanced level training. It is expected that the present manual will fill this gap.
The chapters have been written and the maps prepared by a number of experts on the individual subjects, largely by present and past staff of the Department of Entomology of the London School of Hygiene and Tropical Medicine initially underthe direction of Professor G. Davidson and, later, Professor A. N. Clements. Each chapter is concluded by a number of references, which may provide guidance and suggestions for further reading.
Dr R. Slooff Director, Division of Vector Biology and Control*For the assistance of trainers in illustrating the information provided in the maps of distribution of arthropod-borne diseases and their principal vectors, a set of slides and another of transparencies for overhead projection are soon to be produced and made available against payment. Orders for this material should be sent to:
The Division of Vector Biology and Control, World Health Organization, 1211 Geneva 27, Switzerland.
Pathogens: Plasmodium falciparum Plasmodium malariae Plasmodium ovale Plasmodium vivax Vectors: Mosquitos of the genus Anopheles (Order Diptera, Family Culicidae, Subfamily Anophelinae). Reservoirs: Probably not zoonotic, although the same or similar parasites occur in other primates in South America, Africa and Asia. Occasional cases of simian malaria transmitted to man have been reported (pathogens, e.g. P. cynomolgi, P. knowlesi.)Since the first decades of the present century the worldwide extent of human malaria has receded, especially in temperate areas, as a result of socio-economic progress and through local control programmes, which developed into the global eradication campaign. However, as may be seen from Map 1 (page 16), the disease remains highly endemic in most countries between latitude 30\'a1N and the Tropic of Capricorn in the south with northerly extensions consisting of Turkey, Syria, Iraq, Iran, Afghanistan, Pakistan and China.
The world malaria control situation has been described by Bruce-Chwatt (1985) and official malariological statistics are published regularly in issues of the WHO Weekly Epidemiological Record and annually in the World Health Statistical Quarterly.
The measures used for the control of malaria include those against the parasite through the widespread use of antimalarial drugs for chemotherapy and chemosuppression and those against the anopheline vector. In the latter case, the principal weapon for the control of malaria transmission has been the use of residual insecticides on the interior surfaces of human dwellings. In addition larvicidal measures including biological control have been employed as well as environmental management to reduce breeding places. Personal protection is an additional measure which may in time include the use of antimalarial vaccines.
The success of spraying programmes for the interruption of malaria transmission has depended upon the vulnerability of local vector species of Anopheles in each endemic area. The more endophilic (indoor-resting) vectors such as A. labranchiae in Italy or A. minimus in northern India have not been difficult to control because their survival-rates are insufficient to maintain malaria transmission in areas with adequate insecticidal coverage, i.e. most structures sprayed regularly with residual insecticide.
Unfortunately, however, after years of selection pressure, as a result of the use of pesticides for public health or agriculture purposes, many vector populations have become resistant to the insecticides used. By the end of 1985 a total of 50 anopheline species were recorded as resistant to one or more pesticides, at least 11 of the 50 species being important vectors of malaria (see WHO Technical Report Series No. 737). A number of the vectors are resistant to both organochlorine and organophosphorus insecticides, but some are also resistant to carbamates and pyrethroids. Among these multiresistant vectors are A. albimanus in Mexico and Central America, the A. culicifacies complex in India and Sri Lanka, A. pharoensis in Egypt, A. sacharovi in Turkey and A. stephensi in India and Iran. Further, some vector populations show exophily to an extent that makes it difficult to control malaria transmission by means of vector control based on house spraying alone. Much appears to depend on the environmental conditions and the genetic background of the vector population. This is particularly evident in the cases of A. albimanus in El Salvador, A. arabiensis and A. pharoensis in Africa, A. balabacensis and A. dirus in southeastern Asia, A. farauti in the island of New Guinea and the Solomon Islands, A. sacharovi in Turkey and A. stephensi in Iran. It is increasingly necessary, therefore, to develop integrated vector control programmes involving community participation, together with malaria chemotherapy delivered through primary health care.
Resistance of P. falciparum to chloroquine was first reported at the beginning of the 1 960s and year by year the areas of reported resistance have gradually extended or developed further to much of southeastern Asia and South America, and is currently becoming an increasingly serious problem in Africa. Resistance of P. falciparum to other antimalarial drugs is also increasing in these areas. The resultant problems for malaria treatment make it all the more important to prevent transmission through vector control and by means of personal protection.
Map 2 (page 17) and the associated table 1 (pages 10 and 11 ) follow the malarial epidemiological zones established by Macdonald (1957), in which he attempted to define areas where the epidemiology of malaria is of the same nature throughout. The similarity within the zone covers temperature, rainfall, seasons, physical features of the land and other factors, as well as on the distribution of anopheline vector species. The tactics for vector control activities must be decided with knowledge of the biology, behaviour and degree of susceptibility to insecticides of the particular Anopheles species that are responsible for malaria transmission in each situation. Each of the 12 zones of malaria epidemiology has certain principal vectors (table 1 [a], [b], Map 2) which generally maintain transmission, plus some secondary vectors which augment the amount of transmission but would not sustain the disease cycle efficiently without the principal vectors.
Many vectors of malaria and other diseases belong to groups of sibling species, i.e. morphologically identical species. Such species are reproductively isolated from each other when they occur sympatrically, i.e. they do not normally interbreed where mixed populations occur. Despite the morphological similarity of sibling species, they show ecological and behavioural contrasts which give rise to differences in their vectorial capacity. Thus it is desirable to distinguish between sibling species using genetic methods of identification, e.g. cytotaxonomy or zymotaxonomy (White, 1984).
Much work is currently still being undertaken on Anopheles complexes of sibling species, but at present some of the most important examples are:
The demonstration of the multispecies nature of some traditional taxa now calls for revision of a great deal of the pre-existing taxonomic and epidemiological observations on anophelines. It may be noted in this connection that the Smithsonian Institution's "Mosquito Information Management Project" is a computerized source of systematic, ecological and distributional data (see Faran, M. E. et al., 1984).
INDEX OF MAPS
The maps listed below are presented on pages 16-22.
MAP 1. Epidemiological assessment of the status of malaria, 1987.
MAP 2. The zones of malaria epidemiology.
Maps 3-8 show the range of species distribution and do not necessarily indicate that a particular species acts as a vector throughout the range shown on the map (see the notes below in respect to A. pharoensis - Map 5e and A. farauti - Map 8a).
Principal vectors of malaria in Zone 1 (North American), Zone 2 (Central American), and Zone 3 (South American).
MAP 3a. A. darlingi, A. aquasalis, A. freebomi, A. quadnrnaculatus.
MAP 3b. A. albimanus, A. nuneztovari, A. pseudopunctipennis.
Sources: Frattini, 1962; Darsie & Ward, 1981; Faran & Linthicum, 1981
Principal vectors in Zone 4 (North Eurasian) and Zone 5 (Mediterranean).
MAP 4a. A. messeae and A. superpictus.
MAP 4b. A. atroparvus and A. labranchiae.
MAP 4c. A. sacharovi.
Sources: Bruce-Chwatt & de Zulueta,1980; Macan,1950; Postiglione et al., 1973; White,1978; Zahar, 1974.
Principal vectors in Zone 8 (Indo-lranian).
MAP 4d. A. culicifacies
Sources: Harrison,1980; Kalra,1978.
MAP 4e. A. fluviatilis
Source: Knight & Stone,1977.
MAP 4f. A. pulcherrimus
Sources: Macan,1950; Postiglione et al., 1973.
MAP 4g. A. stephensi
Sources: Macan,1950; Zahar,1974.
Principal vectors in Zone 7 (Afrotropical).
MAPS 5a, 5b, 5c. Principal members of the Anopheles gambiae complex:
5a. A. arabiensis, 5b. A. gambiae, 5c. A. melas and A. merus.
Sources: Coluzzi et al., 1979; White,1985.
MAP 5d. A. funestus
Source: Gillies & de Meillon, 1968.
Principal vectors in Zone 6 (Desert).
MAP 5e. A. pharoensis
This taxon is regarded as an important malaria vector only in Egypt (Zahar, 1974), but at least two sibling species are widespread in Africa (Miles et al., 1983). Perhaps only one of these species is the vector and predominant in Egypt.
Source: Gillies & de Meillon,1968.
MAP 5f. A. sergentii
Source: Farid,1956.
Principal vectors of malaria in Zone 9 (Indo-Chinese Hills), Zone 10 (Malaysian) and Zone 11 (Chinese).
MAP 6a. A. campestris and A. donaldi.
MAP 6b. A. letifer.
MAP 6c. A. nigerrimus.
MAP 6d. A. aconitus.
MAP 6e. A. barbirostris.
MAP 6f. A. balabacensis and A. dirus.
MAP 6g. A. leucosphyrus.
MAP 6h. A. ludlowae and A. maculatus.
MAP 7a. A. minimus and A. flavirostris.
MAP 7b. A. philippinensis.
MAP 7c. A. sundaicus.
MAP 7d. A. sinensis.
Sources: Brown & Pal,1971; Harrison, 1980; Harrison & Scanlon,1975; Kalra,1978; Rao,1984; Reid, 1968, Reid et al., 1979; White,1983.
Principal vectors of malaria in Zone 12 (Australasian): the A. punctulatus complex.
MAP 8a. A. farauti - three sibling species have been distinguished by Bryan (1973) and Mahon & Miethke (1982), provisionally called No. 1 which is the malaria vector in the islands, and Nos. 2 and 3 which are known only from Australia where malaria has been eradicated.
MAP 8b. A. koliensis.
MAP 8c. A. punctulatus.
Source: Belkin,1962.
REFERENCES
Belkin, J. N. The mosquitoes of the South Pacific (Diptera, Culicidae). Berkeley, University of California Press,1962, Vol. l, 608 pp.; Vol. ll, plates 1-412.
Brown, A. W. A. & Pal, R. Insecticide resistance in arthropods. Geneva, WHO Monograph Series, No. 38,1971, 491 pp.
Bnuce-Chwatt, L. J. Essential malariology (2nd ed.). London, Heinemann 1985, xii + 452 pp.
Bnuce-Chwatt, L. J. & de Zulueta, J. The rise and fall of malaria in Europe: a historico-epidemiological study. Oxford, University Press,1980, 240 pp.
Bryan, J. H. Studies on the Anopheles punctulatus complex. Transactions of the Royal Society of Tropical Medicine and Hygiene, 67: 64-91 (1973).
Coluzzi, M., Sabatini, A., Petrarca, V. & Di Deco, M. A. Chromosomal differentiation and adaptation to human environments in the Anopheles gambiae complex. Transactions of the Royal Society of Tropical Medicine and Hygiene, 73: 483-497 (1979) .
Darsie, R. F. & Ward, R. A. Identification and geographical distribution of the mosquitoes of North America, north of Mexico. (Mosquito Systematics Suppl. No.1). Fresno, American Mosquito Control Association,1981, 313 pp.
Faran, M. E. & Linthicum, K. J. A handbook of the Amazonian species of Anopheles (Nyssorhynchus) (Diptera: Culicidae). Mosquito Systematics, 13: 1-81 (1981).
Faran, M. E. et al. A computerized mosquito information and collection management system for systematic research and medical entomology. Mosquito Systematics, 16: 289-307 (1984).
Farid, M. A. The implications of Anopheles sergentii for malaria eradication programmes east of the Mediterranean. Bulletin of the World Health Organization, 15: 821 -828 (1956).
Forattini, O. P. Entomologia Medica, Vols 1 & 2. Sao Paulo, University Press,1962.
Gillies, M . T. & de Meillon, B. The anophelinae of Africa south of the Sahara. Johannesburg, South African Institute for Medical Research, Publication No. 54, 1968, 343 pp.
Harrison, B. A. Medical entomology studies. Xlll. The Myzomyia series of Anopheles (Cellia), with emphasis on intra-interspecific variations (Diptera: Culicidae). Contributions of the American Entomological Institute (Ann Arbor), 17(4): 1 -195 (1980).
Harrison, B. A. & Scanlon, J. E. Medical entomology studies. Il. The subgenus Anopheles in Thailand (Diptera: Culicidae). Contributions of the American Entomological Institute (Ann Arbor), 12: 1 -307 (1975).
Kalra, N. L. National malaria eradication programme, India - its problems, management and research needs. Journal of Communicable Diseases, 10: 1-20 (1978).
Knight, K. L. & Stone, A. A catalogue of the mosquitoes of the world (Diptera: Culicidae) (Thomas Say Foundation, Vol. Vl). College Park, Entomological Society of America, 1977, 610 pp.
Macan, T. T. The anopheline mosquitoes of Iraq and North Persia. pp.109-219 In: Leeson, H. S., ed. Anopheles and Malaria in the Near East (Memoirs of The London School of Hygiene and Tropical Medicine, No. 7). London, Lewis, 1950.
Macdonald, G. The epidemiology and control of malaria. Oxford, University Press, 1957.
Mahon, R. J. & Miethke, P. M. Anopheles farauti No. 3, a hitherto unrecognized biological species of mosquito within the taxon A. farauti Laveran. Transactions of the Royal Society of Tropical Medicine and Hygiene, 76: 8-12 (1982).
Miles, S. J., Green, C. A. & Hunt, R. H. Genetic observations on the taxon Anopheles (Cellia) pharoensis Theobald (Diptera: Culicidae). Journal of Tropical Medicine and Hygiene 86: 153-157 (1983).
Postiglione, M., Tabanli, B. & Ramsdale, C. D. The Anopheles of Turkey. Rivista de Parassitologia, 34: 127-159 (1973).
Rao, T. R . The anophelines of India. New Delhi, Malaria Research Centre, 1984, 518 pp.
Reid, J. A. Anopheline mosquitoes of Malaya and Bomeo. Studies from the Institute for Medical Research, Malaysia. No. 31. Kuala Lumpur, Government of Malaysia, 1968, 520 pp.
Reid, J. A., Harrison, B. A. & Atmosoedjono, S. Variation and vector status in Anopheles barbirostris. Mosquito Systematics, 11: 235-251 (1979).
White, G. B. Systematic reappraisal of the Anopheles maculipennis complex. Mosquito Systematics, 10: 13-44 (1978).
White, G. B. The importance of Anopheles leucosphyrus group mosquitoes as vectors of malaria and filariasis in relation to transmigration and forestry in Indonesia, with assessment of Anopheles balabacensis ecology and vectorial capacity. Unpublished report to WHO-VBCRU, Jakarta, 1983, 55 pp.
White, G. B. Needs and progress in the application of new techniques to mosquito identification. In: Newton, B. N. & Michal, F., eds. New approaches to the identification of parasites and their vectors. Proceedings of a Symposium on Application of Biochemical and Molecular Biology Techniques to Problems of Parasite and Vector Identification. UNDP/World Bank/WHO Special Programme, Geneva. Basle, Schwabe, 1984, pp. 293-332.
White, G. B. Anopheles bwambae sp.n., a malaria vector in the Semliki Valley, Uganda, and its relationships with other sibling species of the Anopheles gambiae complex (Diptera: Culicidae). Systematic Entomology, 10: 501-522 (1985).
WHO Technical Report Series, No. 737, 1986 (Resistance of vectors and reservoirs of disease to pesticides: tenth report of the WHO Expert Committee on Vector Biology and Control). Geneva, World Health Organization, 87 pp.
Zahar, A. R. Review of the ecology of malaria vectors in the WHO Eastern Mediterranean Region. Bulletin of the World Health Organization, 50: 427-440 (1974).
Pathogens: Brugian filariasis Brugia malayi--periodic and subperiodic forms Brugia timori--periodic form Bancroftian filariasis Wuchereria bancrofti--periodic and subperiodic forms Vectors: Many of those Anopheles which are principal malaria vectors (Maps 3-8), together with various species of culicine mosquitos in the genera Aedes, Culex and Mansonia. In urban foci the vectors are invariably Culex quinquefasciatus in tropical countries, or C. pipiens in temperate countries. Whereas Brugia and Wuchereria usually show nocturnal periodicity of microfilariae in the host peripheral blood, which is an adaptation for uptake and transmission by vectors that feed only at night, there are some situations where subperiodic strains of these parasites are adapted for transmission by diurnally active vectors (some Mansonia and Aedes spp.). See Table 2[a], [b], [c], [d], for further information. Reservoirs: Periodic Brugia spp. and Wuchereria bancrofti are found almost exclusively in man. Brugia malayi subperiodic strains are common in domestic cats, various wild carnivores and especially the Macaca and Presbytis monkeys, from which transmission to man is not unusual via diurnally active Mansonia spp.Human lymphatic filariasis occurs in humid tropical foci in Africa, the Americas, Asia, and numerous islands in the Pacific Ocean. Local epidemiology of the disease is strongly influenced by the behaviour and ecology of the various species of mosquito vectors, so that seven principal epidemiological zones of Bancroftian filariasis distribution are based on regional variations of the vectors (Map 9a, page 31). Brugian filariasis is restricted to parts of Southeast Asia and Australasia (Map 9b, page 31).
Unlike malaria or arboviruses, which may be transmitted by the bite of a single infective vector, filarial infections require repeated inoculation of infective larvae - perhaps hundreds per year - in order for the worms to reproduce successfully and produce microfilaraemia. Often the disease is asymptomatic initially with subsequent episodes of acute inflammation of the lymphatic system and fever, with the chronic stage of "elephantiasis" developing only after many years, if at all, by which time the microfilariae have usually disappeared from the bloodstream of the patient. Lymphatic filariasis has been reviewed comprehensively by Sasa (1976) and more recently in WHO Technical Report No. 702 (1984).
Bancroftian filariasis. In urban situations, W. bancrofti is becoming increasingly prevalent in warm countries due to transmission by Culex quinquefasciatus and C. pipiens (Map 10, page 32), which breed prolifically in polluted water. As these mosquitos have developed widespread resistance to insecticides (WHO,1986) it is preferable to control them by environmental management and source reduction (WHO, 1982).
In rural situations, W. bancrofti is mainly transmitted by some of the same Anopheles spp. which are the principal vectors of malaria. The control of Bancroftian filariasis as a side-effect of antimalarial house-spraying has therefore been a widespread bonus in many parts of the world. Such strains of the parasite are nocturnally periodic, but in some areas (e.g. West Africa, Malaysia) they cannot be transmitted by urban Culex quinquefasciatus because these mosquitos are apparently refractory to anopheline-adapted strains of W. bancrofti. The reverse situation applies in other areas (e.g. India, Malaysia) where Culex-adapted strains of W. bancrofti cannot be transmitted by anophelines. In Polynesia (Zone 7), where no anophelines occur, W. bancrofti forms are subperiodic, being adapted to uptake and transmission by the Aedes (Stegomyia) scutellaris group which bite during daytime and evening (Macdonald, 1976). In a few forest situations of Southeast Asia (Zone 6), there are also subperiodic strains of W. bancroftitransmitted by the Aedes (Finlaya) niveus group, which may indicate the possible origins of the Polynesian subperiodic strain.
Brugian filariasis. Brugia malayi occurs only in South Asia (Map 9b), where its distribution and prevalence have been reduced by control of vector Mansonia spp. by the simple method of removal of the host-plants from their breeding-places. Subperiodic strains occur in swamp-forest habitats, transmitted mainly by the M. bonneaedives group as a zoonotic cycle. Periodic strains of B. malayi are transmitted also by Anopheles spp. and are usually not zoonotic (Wharton, 1962). The subperiodic forms are transmitted by the Mansonia spp. and by Coquillettidia crassipes.
Brugia timori is restricted to Indonesian islands of Flores and Timor and of others nearby. It is apparently not zoonotic, and the only known vector is Anopheles barbirostris. It is interesting that B. timori can apparently not be transmitted by various other mosquito species such as A. subpictus which is a vector of both malaria and W. bancrofti among people in that area.
INDEX OF MAPS
The maps listed below are presented on pages 31 - 34.
MAP 9. Distribution of human Iymphatic filarial parasites in the major endemic zones in the world.
MAP 10. Culex pipiens and C. quinquefasciatus, the urban vectors of nocturnally periodic strains of W. bancrofti.
MAP 11. Mansonia uniformis in New Guinea and the Bismarck Archipelago, Aedes (Ochlerotatus) vigilax in islands of the southwest Pacific, vectors of nocturnally periodic strains of W. bancrofti in rural areas; Aedes (Stegomyia) scutellaris group vectors of subperiodic strains of W. bancrofti in Polynesian islands.
MAP 12a. Aedes (Finlaya) niveus group (including A. harinasutai), vector in Thailand of subperiodic W. bancrofti in some forest localities.
MAP 12b. Aedes (Finlaya) poicilius, vector of periodic W. bancrofti in some Philippines islands where people live close to abaca (Musa textiles) plantations and the vector breeds prolifically in axils of the abaca plants.
MAPS l2c-l2f. Mansonia spp. vectors of B. malayi (see Table 2[a], [b], [c], [d], and MAP 9).
MAP 12g. Old world distribution of Mansonia uniformis, the most widespread species of genus Mansonia, a rural vector of periodic W. bancrofti in New Guinea but not elsewhere, also a vector of both periodic and subperiodic strains of B. malayi at least in Malaysia.
Sources: Barraud, 1934; Basio, 1971; Belkin, 1962; Delfinado, 1966; Dobrotworsky, 1965; Huang & Hitchcock, 1980; Knight & Stone, 1977; Leihne et al.,1976; Macdonald, 1976; Sasa, 1976; Tanaka et al.,1979; Wharton, 1962; White, 1982; WHO, 1984.
REFERENCES
Barraud, P.J. The fauna of British India. Diptera, Vol. V. Family Culicidae, Tribes Megarhini and Culicini. London, Taylor and Francis, 1934. 463 pp.
Basio, R.G. The mosquito fauna of the Philippines. National Museum of The Philippines, Monograph No. 4. Manila,1971. 198 pp.
Belkin, J.N. The mosquitoes of the South Pacific (Diptera, Culicidae). Berkeley, University of California Press, 1962, Vol. l, 608 pp; Vol. ll, plates 1-412.
Delfinado, M.D.
Dobrotworsky, N.V. The mosquitoes of Victoria. Melbourne, University Press, 1965. 237 pp.
Huang, Y.-M. & Hitchcock, J.C. A revision of the Aedes scutellaris group of Tonga (Diptera: Culicidae). Contributions of the American Entomologlcal Institute (Ann Arbor), 17: 1-107 (1980).
Knight, K.L. & Stone, A. A catalogue of the mosquitoes of the world (Diptera: Culicidae) (Thomas Say Foundation, Vol. Vl). College Park, Maryland, Entomological Society of America,1977. 611 pp.
Liehne, P.F.S., Stanley, N.F., Alpers, M.P. & Liehne, C G. Ord River arboviruses - the study site and mosquitoes. Australian J. Exp. Biol. Med. Sci., 54: 487-497 (1976)
Macdonald, W.W. Mosquito genetics in relation to filarial infections. pp.1-24 In: Taylor, A.E.R. & Muller, R. (ed.) Genetic aspects of host-parasite relationships (Symposia of the British Society for Parasitology, Vol.14). Oxford, Blackwell,1976.
Sasa, M. Human filariasis. Tokyo, University of Tokyo Press. 1976, 819 pp.
Tanaka, K., Mizusawa, K. & Saugstad, E.S. A revision of the adult and larval mosquitoes of Japan (including the Ryukyu Archipelago and the Ogasawara Islands) and Korea (Diptera: Culicidae). Contributions of the American Entomological Institute (Ann Arbor), 16: 1-98 (1979) .
Wharton, R.H. The biology of Mansonia mosquitoes in relation to the transmission of filariasis in Malaya. Bulletin of the Institute of Medical Research of the Federation of Malaysia, 11: 1 -114 (1962).
White, G.B. Medical entomology: mosquitoes. pp. 1404-1435 In: Manson-Bahr, P.E.C. & Apted, F.l.C. (ed.) Manson's Tropical Diseases, 19th Edition. London, BailliereTindall,1987. 1557 pp.
WHO. Manual on environmental management for mosquito control. Geneva, World Health Organization, Offset Publication No. 66,1982. 283 pp.
WHO Technical Report Series, No. 702,1984. (Lymphatic filariasis: fourth report of the WHO Expert Committee on Filariasis). Geneva, World Health Organization, 112 pp.
WHO Technical Report Series, No. 737, 1986. (Resistance of vectors and reservoirs of disease to pesticides: tenth report of the WHO Expert Committee on Vector Biology and Control). Geneva, World Health Organization, 87 pp.
In East Africa, A. africanus, a rain forest canopy species associated with primate hosts, is involved as well as A. simpsoni which breeds in vegetation around human dwellings. A. simpsoni may act as a link vector associating the forest cycle with a rural/ urban cycle involving A. aegypti, however, A. simpsoni was also believed to have been responsible for man to man transmission during the Ethiopian outbreak in 1960-1962. A. luteocephalus extends from the forest to the savanna and in the savanna may enter villages to bite man. In the New World human infections are almost exclusively from direct contact with exposure to infected forest canopy mosquitos. All the Haemagogus spp., transmitting jungle yellow fever, breed in forest treeholes or occasionally in bamboo stumps.
Experimental transovarial transmission of the yellow fever virus in mosquitos has been demonstrated, and the virus has been recovered from eggs and larvae of the tick Amblyomma variegatum in Central Africa. The disease has never been reported from Asia despite the presence of the vector A. aegypti and considerable populations of susceptible monkeys.
INDEX OF MAPS
The maps listed below are presented on pages 47 - 50.
MAP 13a, b Extent of dengue and dengue haemorrhagic fever outbreaks
MAP 13c Yellow fever endemic areas.
MAP 14 Distribution of Aedes aegypti in 1930 and 1980.
MAP 15 Old World Distribution of Aedes albopictus (15a); and that of A. africanus (15b); A. luteocephalus (15c); A. simpsoni complex (15d); A. furcifer (15e); A. taylori (15f) in Africa.
MAP 16 Distribution of Haemagogus leucocelaenus (16a); H. janthinomys (16b); H. spegazzinii (16c); Sabethes chloropterus (16d) in the Americas.
Dengue haemorrhagic fever is a severe, often fatal disease characterized by fever, shock, acute haemorrhage and high mortality, usually affecting younger children. First described from the Philippines in 1953 (although a similar "syndrome" was noted in Australia and China much earlier), it is now known to occur in many of the urban centres of Southeast Asia and is one of the leading causes for hospitalization and death in children in tropical Asia. In the Caribbean, dengue haemorrhagic fever was reported for the first time in epidemic form in Cuba, in 1981, and an increasing number of sporadic cases are now occurring in that region. It is believed by some experts that this form of the disease is an immunopathological phenomenon due to sequential infections with more than one serotype.
It is possible that dengue was carried to the Mediterranean (from where it has disappeared following the eradication of A. aegypti) from East Africa in the late 19th century, through the slave trade out of Zanzibar and via the Red Sea ports. There is evidence to suggest that dengue originated from foci in tropical Asia and from there spread to Africa. Originally serotypes dengue 1 and 2 were found in West Africa, 2 in East Africa, Seychelles and La Reunion, 3 in Mozambique and 4 in the Pacific. Rapid air travel is believed to have introduced dengue 1 from Africa to the Caribbean and dengue 4 from the Pacific to the Caribbean where outbreaks of both serotypes followed.
The peri-domestic, highly anthropophilic Aedes aegypti is the principal vector of all the serotypes and the sole vector in the New World and Australia. All areas where A. aegypti is present are therefore potentially dengue-risk areas. Aedes albopictus (Map 15a) may be the sole vector in some rural areas of Southeast Asia. A recent outbreak of dengue 2 in the Seychelles was due to A. albopictus, and this vector has now invaded the Americas, spreading to some 16 states in the USA, to Mexico and to four states in Brazil. In the Southwest Pacific, A. aegypti, as well as members of the A. scutellaris complex, are the principal or sole vectors. Dengue haemorrhagic fever is associated always with urban centres and A. aegypti. Until recently man was believed to be the only host for dengue viruses, but Knudsen et al. (1977) and Rudnick (1983), working in Malaysia, discovered a jungle cycle of dengue involving canopy feeding monkeys and forest mosquitos of the Aedes niveus group which feed on man and monkeys.
Transovarial transmission of the virus in A. aegypti and A. albopictus has been demonstrated and the virus has been recovered from field-collected larvae of A. aegypti in Burma, suggesting the occurrence of transovarial transmission in nature.
INDEX OF MAPS
The maps listed below are presented on pages 33, 48 and 49.
MAP 11. Mansonia uniformis in New Guinea and the Bismarck Archipelago, Aedes (Ochlerotatus) vigilax in islands of the southwest Pacific, vectors of nocturnally periodic strains of W. bancrofti in rural areas; Aedes (Stegomyia) scutellaris group vectors of subperiodic strains of W. bancrofti in Polynesian islands.
MAP 14 Distribution of Aedes aegypti in 1930 and 1980.
MAP 15 Old World Distribution of Aedes albopictus (15a); and that of A. africanus (15b); A. luteocephalus (15c); A. simpsoni complex (15d); A. furcifer (15e); A. taylori (15f) in Africa.
The virus is maintained in nature by mosquitos and non-human vertebrates, and man becomes accidentally involved. In temperate countries like Japan, the disease occurs in the warm weather; in the tropics it could occur during any season, although the risk is higher during and immediately after the rainy season when the mosquito population increases. Culex tritaeniorhynchus (Map 17d), a rice field breeding mosquito is the principal vector and feeds mainly on large animals and birds. Elsewhere in the area of distribution, C. gelidus (predominantly a pig biter) (Map 17b) and C. vishnui group (Map 17c) mosquitos are also involved. The virus has been isolated from other Culex spp. as well as Aedes and Anopheles mosquitos.
Japanese encephalitis is predominantly a rural disease and in most of southeastern Asia, associated with rice cultivation and mosquitos which breed in rice fields. Transmission is by bite; transovarial transmission of the virus has been demonstrated in the laboratory in some Aedes species. One vaccine for protection of human beings is commercially available.
INDEX OF MAPS
The maps listed below are presented on page 51.
MAP 17a Extent of Japanese encephalitis endemicity.
MAP 17b Distribution of Culex gelidus.
MAP 17c Distribution of Culex vishnui group.
MAP 17d Distribution of Culex tritaeniorhynchus.
ARBOVIRUSES
More than 400 viruses have been isolated and characterized as being arthropod-borne viruses. In many cases they have been isolated only once or a few times, from pooled specimens of arthropods, and there is no evidence as to what may be their possible vertebrate hosts. Others, such as dengue (DEN), yellow fever (YF) and Japanese encephalitis (JE) have been investigated exhaustively and the vector/host cycles are well known in at least some situations. They are all zoonotic, and almost 100 arboviruses have been recorded as causing clinical symptoms in man. Most are transmitted by vector mosquitos (Table 3) or ticks. Some of the principal diseases caused by arboviruses, and distributions of their most important vectors, are shown in the maps.
Symptoms of acute encephalitis are caused by several groups of arboviruses, notably the California group. In general such arboviruses have a wide range of vertebrate reservoir hosts and many alternative vectors.
In the northeastern USA (Map 18a), sporadic cases of La Crosse virus give cause for concern. The vectors of this virus are the tree-hole breeding pair of sibling species Aedes hendersoni (Map 19b), which breeds higher up the trees and has less contact with man than A. triseriatus (Map 19b), which is the vector of most widespread importance. A. triseriatus has also begun to adapt to breeding in artificial containers, especially old car tyres. Other important vectors of CE Group viruses are A. atlanticus and A. trivittatus (Map 19a) which breed in ground pools and A. canadensis (Map 20c). There is evidence that CE Group viruses are often transmitted transovarially from generation to generation in the vectors.
Alphavirus)
INDEX OF MAPS
The maps listed below are presented on pages 52 - 54.
Distribution of encephalitis epidemics (occasional cases occur elsewhere as the viruses are enzootic, see text):
MAP 18a. California Encephalitis (CE), including La Crosse virus (LAC).
MAP 18b. Saint Louis Encephalitis (SLE).
MAP 18c. Western Equine Encephalitis (WEE).
MAP 18d. Eastern Equine Encephalitis (EEE) and Venezuelan Equine Encephalitis (VEE).
Sources: Theiler & Downs, 1973; Monath, 1979; Sturchler, 1981.
Distribution of principal vectors of major encephalitides in the Americas (Parts I & Il):
MAP 19a. Aedes atlanticus and A. trivittatus, vectors of CE group viruses.
MAP 19b. Aedes hendersoni and A. triseriatus, vectors of LAC.
MAP 19c. Culex tarsalis, vector of SLE and WEE.
MAP 19d. Culiseta melanura, vector of EEE.
MAP 20a Aedes taeniorhynchus, vector of EEE and VEE, in the Americas.
MAP 20b Aedes sollicitans, vector of EEE and VEE, Aedes serratus, vector of VEE.
MAP 20c Aedes canadensis, vector of CE group viruses, Culex nigripalpus, vector of SLE.
MAP 20d Psorophora ferox and Culex portesi, vectors of VEE.
Sources: Darsie & Ward, 1981; Forattini, 1962; Harwood & James, 1979; Knight & Stone, 1977; Sudia & Newhouse, 1975; Watts et al., 1976; White, 1987.
REFERENCES
Arnell, J.H. Mosquito studies (Diptera, Culicidae) XXXII. A revision of the genus Haemagogus. Contributions of the American Entomological Institute, 10(2): 1 - 174 (1973).
Belkin, J.N. The mosquitoes of the South Pacific. Berkeley, University of California Press. Vol. l, 608 pp; Vol. ll, Plates 1-412, 1962.
Brian, G.W., ed. Handbook series in zoonoses, Section B. Viral zoonoses. Vol.1,1981, 510 pp.
Brown, A.W.A. Yellow fever, dengue and dengue haemorrhagic fever. pp. 271-317. In: Howe, G.M. ed. A world geography of human diseases, London, Academic Press, 1977, 621 pp.
Calisher, C.H. & Thompson, W.H. eds. California serogroup viruses. New York, Liss, 1983.
Christophers, S.R. Aedes aegypti, the yellow fever mosquito. Cambridge, University Press, 1960, 739 pp.
Darsie, R.F. & Ward, R.A. Identification and geographical distribution of the mosquitoes of North America, north of Mexico (Mosquito Systematics Suppl.1). Fresno, California, American Mosquito Control Association, 1981, 313 pp.
Delfinado, M.D. & Hardy, D.E. A catalogue of the Diptera of the Oriental Region. Vol. I. Nematocera. Honolulu, University Press of Hawaii, 1973, 618 pp.
Euchler, D. Endemiegebiete tropischer Infektionskrankheiten: Karten und Texte fuer die Praxis. Berne, Hans Huber,1981, 246 pp.
Forattini, O.P. Entomologia Medica, Vol. l & Il. Sao Paulo, University Press, 1962.
Forattini, O.P. Entomologia Medica. Vol. III. Culicini. Sao Paulo, University Press, 1965, 416 pp.
Halstead, S.B. Dengue haemorrhagic fever - a public health problem and a field for research. Bulletin of the World Health Organization, 58: 1-21 (1980).
Harwood, R . F. & James, M .T. Entomology in human and animal health. New York, MacMillan, 1979, 548 pp.
Huang, Y.M. Contributions to the mosquito fauna of Southeast Asia. XIV. The subgenus Stegomyia of Aedes in Southeast Asia. I. The Scutellaris group of species. Contributions of the American Entomological Institute, 9 (1): 1-109 (1972).
Huang, Y.M. Medical entomology studies Xl. The subgenus Stegomyia of Aedes in the Oriental Region with keys to the species (Diptera: Culicidae). Contributions of the American Entomological Institute 15 (6): 1-79 (1979).
Huang, Y.M. Aedes (Stegomyia) simpsoni complex in the Ethiopian Region with lectotype designation for simpsoni (Theobald) (Diptera: Culicidae). Mosquito systematics, 11: 221-234 (1979)
Karabatsos, N. ed. International catalogue of arboviruses, including certain other viruses of vertebrates. 3rd Ed. Baltimore, American Society of Tropical Medicine and Hygiene,1985,1147 pp.
Khin, M.M. & Thraw, K.A. Transovarial transmission of dengue 2 viruses by Aedes aegypti in nature. American Journal of Tropical Medicine and Hygiene, 32: 590-594 (1983).
Knight, K.L. & Stone, A. A catalogue of the mosquitoes of the world (Diptera: Culicidae). (Thomas Say Foundation Vol. Vl). College Park M.D. Entomological Society of America,1977, 611 pp.
Knudsen, A.B. The silent jungle transmission cycle of dengue virus and its tenable relationship to endemic dengue in Malaysia. The Malayan Nature Journal, 31: 41-47 (1977).
Leake, C.J. Strategies for vector-borne disease control in rice production systems in developing countries: arboviruses other than Japanese encephalitis. In: Vector-bome disease control in humans through rice agroecosystem management. International Rice Research Publication, in collaboration with the WHO/FAO/UNEP Panel of Experts on Environmental Managemen for Vector Control,1988, pp.161 173.
Macdonald, W.W. Mosquito genetics in relation to filarial infections. pp. 1-24 In: Taylor A.E.R. & Muller, R. ed. Genetic Aspects of host-parasite relationships. (Symposium of the British Society for Parasitology, Vol. 14). Oxford, Blackwells, 1976, pp. 1-24.
Mattingly, P.F. The sub-genus Stegomyia (Diptera: Culicidae) in the Ethiopian Region. Bulletin of the British Museum (Natural History), Entomology, 2 (5): 235-304 ( Part 1) (1952).
Monath, T.P. Arthropod-borne encephalitides in the Americas. Bulletin of the World Health Organization, 57: 513-533 (1979)
Monath, T.P. St. Louis encephalitis. Washington DC, American Public Health Publication,1980.
PAHO. Dengue outbreak in Boa Vista, Roraima Territory, Brazil. Epidemiological Bulletin, 3 (6): 5-7 (1982).
Reinert, J.F. The zoogeography of Aedes (Diceromyia), - Theobald, (Diptera: Culicidae). Journal of the Entomological Society of southern Africa 33: 129-141 (1970).
Rosen, L. et al. Transovarial transmission of dengue viruses by mosquitoes: Aedes albopictus and Aedes aegypti. American Journal of Tropical Medicine and Hygiene, 32, 1108-1119 (1983).
Rudnick, A. The ecology of the dengue virus complex in Peninsular Malaysia. In: Pang, T. & Pathmanathan, R. Proceedings of the International Congress on Dengue/Dengue Haemorrhagic Fever, Kuala Lumpur, University of Malaya, 1983, pp. 7-14.
Sudia, W.D. & Newhouse, V.F. Epidemic Venezuelan Equine Encephalitis in North America: a summary of virus-vector-host relationships. American Journal of Epidemiology, 101: 1-13, (1975).
Tanaka, K. Mizusawa, K., Saugstad, E.S. A revision of the adult and larval mosquitoes of Japan (including the Ryukyu Archipelago and the Ogasawara Islands) and Korea (Diptera: Culicidae). Contributions of the American Entomological Institute, 16: 1-987 (1979).
Watts, D.M., De Foliart, G.R., Yuill, T.M. Experimental transmission of Trivittatus Virus (California Virus group) by Aedes trivittatus. American Journal of Tropical Medicine and Hygiene, 25: 173-176 (1976).
White, G.B. Medical entomology: mosquitoes. In: Manson-Bahr, P.E.C. & Bell, D.R. eds. Manson's Tropical Diseases, 19th Ed. London, Bailliere Tindall, 1987, pp. 1404-1435.
WHO. Yellow Fever in 1979. Weekly Epidemiological Record, 55: 345-351 (1980).
WHO. Overall review of yellow fever in Africa and America, 1965-1979. Weekly Epidemiological Record, 55:356-360 (1980).
WHO. Status of the Aedes aegypti eradication campaign in the Americas: Correction. Weekly Epidemiological Record, 55: 391 (1980).
WHO. Dengue surveillance. Weekly Epidemiological Record, 56: 237-239 (1981).
WHO. Yellow fever in 1981. Weekly Epidemiological Record, 57: 297-301 (1982).
Zavortink, T.J. (1972). Mosquito studies (Diptera, Culicidae) XXVIII. The New World species formerly placed in Aedes (Finlaya). Contributions of the American Entomological Institute, 8: (3): 1 -206 (1972).
Transmission to man is usually by bite of adult infected ticks. The virus is passed trans-stadially in ticks, but there is no evidence of transovarial transmission and the long (overwinter) persistence of the virus in ticks and rodents account for its maintenance in nature. In humid mountainous areas where rodents and ticks are numerous, the incidence of the infection in D. andersoni is 10% or higher.
During the 1960s, up to 80% of RSSE cases were in residents within a 3-8 km radius of their homes, contracted during holidays and weekend trips to the forests, a major pleasure for city-bound people. The reported "spread" of TBE in western Europe is almost certainly due to increased man-tick contact following an increase in leisure and sporting activities which bring man into infected biotopes.
Wandering and migrating birds may spread the ticks into new localities. The viruses are maintained in their tick vectors by trans-stadial passage and by transovarial passage to succeeding generations.
The most abundant tick species in the enzootic area is Haemaphysalis spinigera, the major vector responsible for epizootics and epidemics of the disease. H. spinigera is widely distributed in the tropical evergreen/deciduous forests of southern and central India and Sri Lanka. KFD virus has been isolated from seven other species of Haemaphysalis as well as from Dermacentor and Ixodes ticks.
Larvae and nymphs of Haemaphysalis spinigera parasitize several species of small forest mammals and birds as well as monkeys. Cattle are heavily parasitized by adults of H. spinigera and help to amplify the tick population. Neither cattle nor birds are thought to have any significant role in virus maintenance, apart from acting as hosts for the vector ticks. Man is parasitized almost exclusively by larvae and nymphs of H. spinigera and in the absence of natural transovarial transmission of the virus in this species, transmission to man is by bite of infected nymphal ticks. Other Haemaphysalis spp., particularly H. turturis maintain the enzootic cycle.
Most human cases occur during the dry months of March to May, following months of peak nymphal tick activity. Since the original outbreak of 1957, outbreaks in other forested areas, all in southern India have occurred, almost certainly due to deforestation/exploitation and the entry of man into already existing foci of infection.
INDEX OF MAPS
The maps listed below are presented on pages 64 and 65.
MAP 21 Distribution of Colorado Tick fever and of Dermacentor andersoni
MAP 22 Distribution of louping ill, tick-borne encephalitis, Russian Spring-Summer encephalitis and Kyasaur Forest disease and their main vectors.
Foci of infection are usually restricted and include huts, caves, stables, piggeries, other animal shelters, resort cabins. Trans-stadial and the high rate of transovarial transmission and the ability of the vector ticks to starve for long periods lead to perpetuation of natural foci in the absence of vertebrate hosts.
Rodents probably act as amplifiers rather than as reservoirs of the infection. Following an infecting feed, the ticks have a disseminated infection of practically all the internal organs. Transmission usually through salivary secretion containing the pathogens, and through contamination of the bite wound through infective coxal fluid produced by feeding ticks just before detachment. Vector species which do not produce coxal fluid while on the host, transmit solely by bite. In contrast to ixodid ticks, the argasid ticks (Ornithodoros) are rapid feeders.
In man the disease is manifested by a dermatosis with a subsequent arthritis, neuropathological manifestations and cardiac involvement. Antibiotics in large doses are curative.
In Europe I. ricinus is widespread with about 100 mammalian, avian and reptilian hosts, living in forest environments under 1200 m altitude.
There is some doubt whether there are some differences in Borrelia burgdofferi in Europe and that in USA though they are immunologically closely similar with shared antigenic determinants.
INDEX OF MAPS
The maps listed below are presented on pages 66 - 68.
MAP 23 Old World distribution of endemic tick-borne relapsing fever and the principal Ornithodoros vectors.
MAP 24 New World distribution of endemic tick-borne relapsing fever and the principal Ornithodoros vectors.
MAP 25 Distribution of erythema borreliosis migrans (Lyme disease) and its major vectors.
Vectors: Ticks of the Family Ixodidae; Dermacentor andersoni
(the Rocky Mountain wood tick), D. variabilis (the American dog
tick), Amblyomma cajennense (the Cayenne tick).
Reservoirs: Primarily ticks, transovarial transmission common;
also small mammals, especially rodents.
Immature stages of Dermacentor andersoni and D. variabilis feed on small mammals; D. andersoni adults attack large wild and domestic herbivores and D. variabilis prefers wild and domestic carnivores; both species feed secondarily on other mammals, including man.
Any stage of a tick vector can acquire the infection by feeding. There is transstadial and transovarial transmission of the pathogen in ticks. Transmission to mammals is by the bite of infected ticks; in the case of infected ticks which have overwintered, it may require a day or two for the pathogen to be reactivated and for transmission to take place. Heavy infection in ticks may sometimes be fatal for engorged female vector ticks and may reduce oviposition and egg development.
The chief vectors are Amblyomma hebraeum (in the South African veld) and Rhipicephalus sanguineus. In France, the disease is so closely associated with rabbits that the incidence dropped considerably following decimation of the rabbit population by myxomatosis. There is transovarial transmission and transmission to man is by tick bite. Infection may also occur by contact of the hands with the skin or eyes after crushing ticks removed from dogs. Urban cases appear to be associated with dogs and dog ticks.
Siberian tick typhus has a wide distribution from the far eastern Pacific through northern and southern Siberia, Mongolia, Kazakh and Kirgiz Republics to the Armenian Republic of the USSR. The disease was first recognized in the 1930s when virgin steppe was extensively colonized. Nine tick species have been reported to be reservoirs and vectors of R. sibirica, the most important being Dermacentor marginatus, D. silvarum, Haemaphysalis concinna and Rhipicephalus sanguineus, the last named being the vector in the western part of the distribution of the disease. There is transstadial and transovarial passage of the agent in ticks and transmission to man is by tick bite.
Queensland tick typhus is related to the rickettsialpox spotted fever caused by R. akari transmitted by mites. It is restricted to dense forest interspersed with grassy savanna or secondary scrub. The vector Ixodes holocyclus is widely distributed in coastal and densely forested areas of Australia and is an unusually indiscriminate feeder, causing tick paralysis. There is some evidence that the tick can transmit the infection by bite within a matter of hours after attachment.
INDEX OF MAPS
The maps listed below are presented on pages 69 and 70.
MAP 26 Distribution of tick-borne typhus fevers (Rocky Mountain Spotted Fever) and their vectors in the Americas.
MAP 27 Distribution of tick-borne typhus fevers of the Old World and their vectors.
REFERENCES
Ackermann, R. Erythema migrans borreliosis in Europe: clinical picture, diagnosis, therapy. IXth International Congress of Infectious and Parasitic Diseases, Munich, July 20-26, 1986. Abstract 1210. Munich, Futuramed.
Aeschlimann, A. Ixodes ricini, Linn,1758 (Ixodoidea: Ixodidae). Essai preliminaire de synthase sur la biologie de cette espece en Suisse. Acta Tropica, 29: 321-340 (1972).
Anastos, G. The ticks, or ixodides, of the USSR, . Washington DC, US Public Health Service, 1957, 397 pp.
Blaskovic, D. Studies on tick-borne encephalitis. Bulletin of the World Health Organization, 36 (suppl. 1),1-94 (1967).
Burgdorfer, W. A review of Rocky Mountain spotted fever (tick-borne typhus), its agent, and its tick vectors in the United States. Journal of Medical Entomology, 12: 269-278 (1975).
Burgdorfer, W. The epidemiology of the relapsing fevers. In: Johnson, R. C., ed. "The biology of parasitic spirochetes." New York, Academic Press,1976, pp.191-200.
Burgdorfer, W. Ecological and epidemiological considerations of the Lyme disease spirochaete, Borrelia burgdorfersi. In: Proceedings IXth International Congress of Infectious and Parasitic Diseases, Munich, July 20-26,1986. Vol 11, 11-15, Munich, MMV Medizin Verlag,1986.
Cooley, R.A. The Rocky Mountain wood tick. Montana State College Experimental Station Bulletin, No. 268, (1932) 58 pp.
Cooley, R.A. The genera Dermacentor and Otocentor (lxodidae) in the United States, with studies in variation. U.S. National Institute of Health Bulletin, No. 171, 1938, 89 pp.
Goubau, P. F. Relapsing fevers. A review. Annales de la Societe Belge de Medecine Tropicale, 64: 335-384 (1984).
Haworth, J. Erythema chronicum migrans. Travel Medicine Intemational, 4: 69-71 (1986).
Hoogstraal, H. Ticks in relation to human diseases caused by viruses. Annual Review of Entomology, 11: 261-305 (1966).
Hoogstraal, H. Ticks in relation to human diseases caused by Rickettsia species. Annual Review of Entomology, 12, 377-420 (1967).
Hoogstraal, H. Ticks and spirochetes. Acta Tropica, 36: 133-136 (1979).
Indian Journal of Medical Research, 56 (4). April (Supplement). Eleven papers on Kyanasur Forest Disease, pp. 497-636 (1968).
Johnson, R. C., Schmid, G. P., Hyde, F. W., Steigerwalt, A. G. & Bremner, D. J. Borrelia burgdorferi sp. nov. Etiologic agent of Lyme disease. International Journal of Systemic Bacteriology, 34: 496-497 (1984) .
Leeson, H. S. Some notes on the recorded distribution of Old World species of Ornithodoros. Bulletin of Entomological Research, 44: 517-526 (1953).
Leeson, H. S. Further notes on the geographical distribution of Old World species of Ornithodoros. Bulletin of Entomological Research, 46: 747-748 (1956) .
Nosek, J. and Blaskovic, D. Ticks as vectors of tick-borne encephalitis (TBE) virus in Europe. In: Proceedings of Ill lnternational Congress on Acarology, Prague, pp. 589-591,1971.
Obasi, O. E. Erythema chronicum migrans. British Journal of Dermatology, 97: 459 (1977).
Petrischeva, P. A. & Skrynnik, A. N. Tick-bome relapsing fever. In: "Geography of natural focal diseases of man". Moscow, Medgiz, 1969, pp. 95-119 [In Russian].
Roberts, F. S.H . Australian ticks. Melbourne, CSI RO,1970, 267 pp.
Rufli, Th. "Lyme disease" der neuen Welt versus Erythema Chronicum Migrans der altem Welt und seine Komplikationen. Zentralblatt Hant. und Geschlech, Krankheiten, 150: 909-914 (1985).
Scrimenti, R. J. Erythemachronicum migrans. Archives of Dermatology, 102: 104-105 (1970).
Steere, A. C., Broderick, T. F. & Malawesta, S. E. Erythema chronicum migrans and Lyme arthritis: epidemiologic evidence of a tick vector. American Joumal of Epidemiology, 108: 312-321 (1978).
Stewart, A., Glass, A., Patel, A., Watt, G., Cripps, A. & Clancy, R. Lyme arthritis in Hunter Valley. Medical Journal of Australia, 1: 139 (1982).
Varma, M.G.R. The acarology of Louping III. Acarologia, fasc. H.S., pp. 241-254 (1964)
Varma, M.G.R. The distribution of Ixodes ricinus in Britain in relation to climate and vegetation. In: Theoretical questions of natural foci of diseases. Proceedings of a Symposium held in Prague,1963. Prague Publishing House of the Czech Academy of Sciences,1965.
Work, T.H. and Tropido, H. Kyanasur Forest Disease: a new infection of man and monkeys in tropical India by a virus of the Russian Spring-Summer Complex. In: Proceedings of the ninth Pacific Science Congress, 17. pp.80-84 (1957).
Larvae of the vector mites, chiggers (the only parasitic stage in the life cycle of the mites), acquire the infection by feeding on infected rodents. The rickettsiae are trans-stadially transmitted through non-parasitic nymphal and adult mites which are predatory on soil arthropods, and transovarially through the eggs to parasitic larvae of the next generation, which transmit the pathogen during feeding on animals and man. For most, but not all human cases, a lesion or "eschar" develops at the site of the infective chigger bite.
INDEX OF MAPS
The map listed below is presented on page 73.
MAP 28 Areas of endemic chigger-borne rickettsiosis and distribution of the Leptotrombidium vectors.
REFERENCES
Audy, J.R. A summary topographical account of scrub-typhus 1908-1946. Bulletin of the Institute of Medical Fesearch, Federation of Malaya, 1: 1-82 (1949).
Blake, F.G. et al. Studies on Tsutsugamushi disease (scrub typhus, mite-borne typhus) in New Guinea and adjacent islands. American Journal of Hygiene, 41: 293-373 (1945).
Tamiya, T. Recent advance in studies of Tsutsugamushi disease in Japan. Tokyo, Medical Culture Inc., 1962, 309 pp.
Traub, R. & Wisseman, Jr. C.L. Ecological considerations in scrub typhus. 1. Emerging concepts. Bulletin of the World Health Organization, 39: 209-218 (1968).
Traub, R. & Wisseman, Jr. C.L. The ecology of chigger-borne rickettsiosis (scrub typhus). Joumal of Medical Entomology, 11: 237-303 (1974).
Historically, louse-borne typhus has been characteristic of impoverished communities, of refugees, and of those affected by war or natural disasters. Typically, it is a disease of temperate rather than warm countries, although mountainous areas in the tropics, or warm countries where the nights are cold, may also be high risk areas. At present, louse-borne typhus is reported mostly from certain mountainous areas in East and Central Africa and in some high Andes communities in South America (Map 29). Symptomless carriers still exist amongst the older age groups in many parts of the world, especially in Europe.
Louse-borne relapsing fever is likewise theoretically cosmopolitan (Map 30), but is particularly prevalent in the Ethiopian region. In the world in general, and Africa in particular, it appears to be again increasing in incidence.
Trench fever is now very rare although it can be found in Central America and there appears to have been a recent minor resurgence in Europe.
Louse-borne rickettsial diseases are transmitted by contamination with the faeces of infected lice when these are nubbed into scratches or abrasions on the skin, are breathed in or even blown into the eye. Faeces dry to a fine powder which may remain infective for three months. Louse bites do not transmit diseases to man, it is the faeces which are dangerous. Nor is it necessary to be louse-infested oneself, mere association with the louse-infested individuals can be enough.
In louse-borne relapsing fever, the body lice take up spirochaetes from the blood of infected persons on whom they are especially prevalent during bouts of fever. The spirochaetes can be found in the louse's stomach for a few hours, but nearly all are digested. Sometimes, however, a few persist and appear about a week later in the insect's body cavity. They appear to do the louse no harm. Infection can only occur if a man crushes an infected louse and releases the spirochaetes into a scratch or mucous membrane. This happens frequently among chronically louse-infested individuals, who may destroy some of their lice by bursting them between their finger nails.
Symptomless carriers of typhus and trench fever constitute the reservoirs of the diseases. Murine or flea-borne typhus is a zoonosis, but there is no certain zoonotic reservoir of louse-borne relapsing fever.
INDEX OF MAPS
The maps listed below are presented on pages 76 and 77.
MAP 29 Endemic foci of louse-borne typhus
MAP 30 Endemic areas of louse-borne relapsing fever.
REFERENCES
Alexander, J. O. Arthropods and human skin. Berlin and New York, Springer-Verlag,1984.
Bryceson, A.D. Quarterly Journal of Medicine, 39: 129-170 (1970).
Farhang-Azad, A. et al., Journal of Medical Entomology, 21: 675-680 (1984)
Farhang-Azad, A. et al., Science 227: 543-545 (1985)
Maunder, J. W. The appreciation of lice. Proceedings of the Royal Institute of Great Britain, 55: 1-32 (1983).
Traub, R., Wisseman, C.L. & Farhang-Azad, A. Tropical Diseases Bulletin 75: 237-317, (1978).
Weidhaas, D.E. & Gratz, N.G., World Health Organization, document WHO/VBC/82.858.
Wilcocks, C . Tropical Diseases Bulletin, 56: 1001 -1006 (1959).
Trypanosoma belong to the class of zoomastigophorea and the causative organisms of sleeping sickness are in the subgenus Trypanozoon of the Salivaria group.
Tsetse flies are now restricted to continental Africa (that is, excluding Madagascar, but including some of the inshore islands, for example Zanzibar), between latitude 15N and 30S. About half of Africa is infested: some 10.4 million square kilometres in all.
Glossina species are large (6-15 mm), narrow-bodied flies, brownish or greyish in colour, with a stout proboscis that projects well forward in front of the head.
The genus includes about 30 species and subspecies. Most of these are not associated with transmission of sleeping sickness, although many transmit animal trypanosomiasis to game and domestic livestock. Members of the genus Glossina are divided conveniently into three groups which are often given subgeneric status. These divisions are: (1) the fusca group (=subgenus Austenina), (2) the palpalis group (= subgenus Nemorhina) (Map 31b) and (3) the morsitans groups (= subgenus Glossina s.s.) (Map 31b). Based originally on small differences in the complex male and female genitalia, this division into three groups is broadly supported by chromosome and enzyme-electrophoretic studies.
The fusca group has 12 species, two of which are divided into several subspecies. Some members of the fusca group are minor vectors of animal trypanosomiasis but none are associated with human disease.
The palpalis group consists of five species (two of these are divided into subspecies). Whilst the palpalis group are also forest-dwellers, the species are found primarily along the shores of lakes and rivers, closely associated with the dense woody vegetation along their margin, along which flies extend far out into the savanna woodlands beyond the boundaries of the forest zones; however, none of the palpalis flies occur along the river systems which drain into the Indian Ocean. Three species are major vectors of sleeping sickness, namely, G. fuscipes (Map 32a), G. palpalis (Map 31c) and G. tachinoides (Map 31d).
G. palpalis consists of two subspecies: G. p. gambiensis west of the Togo-Benin savanna gap and G. p. palpalis eastward to Cameroon and south to Angola. Intermediate forms are sometimes found in the area where the subspecies overlap. G. fuscipes and its subspecies are closely related to G. palpalis and intermediate forms occur in the areas of overlap between G. p. palpalis and G. f. fuscipes (Cameroon, Congo and Gabon).
The subspecies of G. fuscipes are: G. f. fuscipes, G. f. martinii and G. f. quanzensis (Map 32a). Together with G. tachinoides, both subspecies of G. palpalis and all three of G . fuscipes are efficient vectors of sleeping sickness; they are characteristically associated with the transmission of the gambiense (chronic) form of the disease typical of western and central Africa. However, both G. f. fuscipes along the shores of Lake Victoria) and G. tachinoides (in the western lowlands of Ethiopia) sometimes also transmit the acute rhodesiense type of disease.
The morsitans group of tsetse (subgenus Glossina) are often referred to as the "savanna" or "game" species, terms which reflect well their very different habitat and distribution, as compared with that of the fusca and palpalis groups considered above. The morsitans flies occupy a huge, non-linear, habitat in the vast areas of bushland and thicket of the drier areas of sub-Saharan Africa: northwards, their limits are the hot, dry conditions near the desert margins (Maps 31b, 32b). To the South, the cold winter climate of the southerly latitudes is a limiting factor. Elsewhere, their distribution may be interrupted by the absence of suitable woody vegetation, and sometimes by the scarcity of the wild pigs and other game animals on which they mainly feed.
G. longipalpis and G. pallidipes are closely-related species with only minor morphological differences, but there is a gap of about 1000 km which separates their areas of distribution. G. longipalpis has no known medical importance, but is an occasional vector of trypanosomes to livestock (Map 32c); G. pallidipes is an important vector of animaltrypanosomiasis throughout its range and has been shown to be a vector of human trypanosomiasis, particularly in the area of Lake Victoria. G. pallidipes is found in dry bushland and thicket vegetation, from Ethiopia and Somalia southwards to Mozambique and Zambia (Map 32c). G. austeni (Map 32d) is clearly related to G. Iongipalpis and G. pallidipes; the three species can be thought of as a species-complex. G. austeni has no known medical importance.
G. swynnertoni, a member of the morsitans group closely related to the G. morsitans subspecies, has a very limited area of distribution, to the south-east of Lake Victoria (Map 32d). The species is capable of transmitting human trypanosomiasis.
As vectors of human (and animal) trypanosomiasis, the subspecies of Glossina morsitans are the most widespread (Map 32b). The recognized subspecies are: G. m. submorsitans, in a broken belt from west Africa across to southern Sudan; G. m. centralis, found in north-central Angola, along the borders of Zambia and Zimbabwe, and in a broad belt running southwestwards from Lake Victoria through to central Zambia. G. m. morsitans is found to the east of the G. m. centralis belt, almost to the Indian Ocean. An eastern form of G. m. submorsitans, ugandensis Vanderplank, 1949, is here given tentative subspecific status (as G. m. ugandensis), since tsetse from G. morsitans populations in western Ethiopia can be consistently ascribed to ugandensis (and distinguished from G. m. submorsitans), by differences in the detailed morphology of the male genitalia. The G. m. ugandensis population occurred at altitudes up to 2000 metres.
INDEX OF MAPS
The maps listed below are presented on pages 85 and 86.
MAP 31a. Distribution of sleeping sickness foci in man in Africa.
MAP 31b. Distribution of Glossina morsitans and G. palpalis groups.
MAP 31c. Distribution of G. palpalis.
MAP 31d. Distribution of G. tachinoides.
MAP 32a. Distribution of G. fuscipes group.
MAP 32b. Distribution of G. morsitans complex.
MAP 32c. Distribution of G. Iongipalpis and G. pallidipes.
MAP 32d. Distribution of G. austeni and G. swynnertoni.
Originally a zoonosis, T. cruzi infection exists in many areas as a silvatic cycle between silvatic Triatominae and small mammals such as rodents and marsupials. In many areas there is no human contact with the silvatic cycle; human infection occurs only through accidental contact with that cycle, or by initiation of a domestic cycle of transmission when bugs invade and colonize houses. Thus the geographical distribution of Chagas disease is less than the distribution of the vectors and silvatic foci of T. cruzi. For example in the southwestern USA, silvatic T. cruzi is widely maintained in woodrat populations by silvatic bug species such as Triatoma protracta; however, domestic cycles of transmission do not occur and only three cases of human Chagas disease have been recorded from the USA, caused by accidental contact with the silvatic cycle. The most complete general work on the subject is the monograph by Lent & Wygodzinsky (1979).
In 1960, WHO estimated that about 7 million people had Chagas disease in Latin America. More recently, Zeledon & Rabinovich (1981 ) estimated that 13-14 million people were infected with T. cruzi, but others have estimated 19-20 million cases on the basis of published serology of blood-donors. Latest data available to WHO indicate some 16 millions infected
or at least serologically positive for T. cruzi and 65 million people are exposed to the risk of infection (PAHO, 1984).
Trypanosoma cruzi, belonging to the Stercolarian group of Trypanosoma is transmitted to man in the faeces of its Triatomine bug vectors, not in the bite. Thus, when an infected bug feeds, it may defecate at the same time. Trypanosomes in the faeces can then penetrate mucous membranes or any wound or abrasion on the skin. Local multiplication of the parasites at the site of entry is followed by blood parasitaemia and intracellular proliferation in diverse tissues, especially muscle. Another important mode of transmission is through blood transfusion from apparently healthy donors in the latent phase of the disease. Transmission via blood t
ransfusion is mainly responsible for the occurrence of Chagas disease in urban areas.
Chagas disease is characterized by an initial acute phase, with high parasitaemia often accompanied by fever, generalized adenopathy, moderate splenomegaly and electrocardiogram changes. The acute phase, which may be symptomless, is followed after 2-3 weeks by the chronic phase with subpatent parasitaemia, which may last up to 15-20 years. During the chronic phase, the parasites penetrate cells of the vital organs, often causing irreversible tissue damage particularly to the autonomic nervous system. Both the early part of the chronic phase, and sometimes the later part, may be symptomless. Death may result from cardiac insufficiency, acute cardiac arrest, or intestinal distortion. The geographical distribution of different clinical pictures of chronic Chagas' disease seems to be matched to some extent with different enzyme patterns of the strains of T. cruzi involved. It is possible that strain differences may contribute to different results from chemotherapy trials in different areas (Miles et al., 1981).
Of the 115 species of Triatominae now recognized, over half have been naturally or experimentally infected with T. cruzi. It seems likely that all species of Triatominae should be regarded as potential vectors. There is no doubt however that some species are more efficient than others. Three factors are of prime importance in vector efficiency: (1 ) degree of susceptibility to T. cruzi infection, (2) time interval between feeding and defecation, (3) degree of contact with human hosts.
The most important vectors, which are highly domestic and build up large colonies in houses, are highly susceptible to T. cruzi infection, and which defecate on the host during or soon after feeding, are as follows:
Tribe: Triatomini
Triatoma infestans (Klug). Widespread in Argentina, Bolivia, Brazil, Chile, Paraguay, Peru and Uruguay. Highly domestic species, not known to colonize silvatic habitats. Geographical distribution expanding (Map 34).
Panstrongylus megistus Burmeister. Widespread in Brazil, also in northeastern Argentina. It has been replaced as a domestic vector by T. infestans in many drier areas, but persists in silvatic foci, sometimes reinvading houses when T. infestans populations are reduced by insecticide application (Map 35).
Triatoma brasitiensis Neiva. Confined to arid zones of northeastern Brazil, especially Ceara, Piaui and Rio Grande do Norte (Map 36).
Triatoma dimidiata Latreille. An important vector in the extreme north of Peru, Ecuador, Colombia, Panama, Costa Rica, Central America into southern Mexico (Map 37).
Tribe: Rhodniini
Rhodnius prolixus Stal. Widespread in Colombia, Venezuela, and possibly in parts of French Guyana, Guyana and Suriname, although there are no recent records from these latter three countries. R. prolixus also occurs in parts of Central America, Guatemala, Honduras, etc., but not in Costa Rica or Panama. It maintains silvatic ecotopes, particularly in birds nests in palm tree crowns (Map 38).
Map 33 shows the approximate distribution of human infection with Trypanosoma cruzi. Note that in the Amazon forest, domestic vectors of Chagas disease are unknown (although silvatic Triatominae are present), and the infection is rare. Human infection is also rare in densely forested parts of Central America.
INDEX OF MAPS
The maps listed below are presented on pages 87 - 92.
MAP 33. Distribution of human infection with Trypanosoma cruzi.
MAP 34. Distribution of Triatoma infestans.
MAP 35. Distribution of Panstrongylus megistus.
MAP 36. Distribution of Triatoma brasiliensis.
MAP 37. Distribution of Triatoma dimidiata.
MAP 38. Distribution of Rhodnius prolixus.
REFERENCES
African trypanosomiasis
Ford, J. The geographical distribution of Glossina. In: Mulligan, H. W. (Ed.). The African trypanosomiases. London, Allen & Unwin, 1970, pp. 274-297.
Jordan, A. M. Systematics. In: M. Laird (Ed.) Tsetse: the future for biological methods in integrated control. Ottawa, International Development Research Centre, 1977, pp.13-22.
Machado, A, de B. Les races geographiques de Glossina morsitans. In: Azevedo, J. F. de (Ed.). First international symposium on tsetse fly breeding under laboratory conditions and its practical applications. Lisbon, Junta de Investigacoes do Ultramar, 1970, pp. 471-486.
Minter, D. M. Tsetse flies (Glossina). In: Manson-Bahr, P. E. C. & Apted, F. I. C. (Eds.) Manson's Tropical Diseases, 19th ed ., London, Bailliere Tindall,1987, pp .1447-1461.
Pollock, J. N. (Ed.) Training Manual for Tsetse Control Personnel, vol.1, Tsetse biology, systematics and distribution. Rome, Food and Agriculture Organization of the United Nations,1982, 280 pp.
de Raadt, P. (1984) African trypanosomiasis. Medicine International, 2(4): 146-150 (1984).
WHO Technical Report Series, No. 635,1979. (The African trypanosomiases: report of a Joint WHO Expert Committee and FAO Expert Consultation), 96 pp.
WHO Technical Report Series No. 739,1986. (Epidemiology and control of African trypanosomiasis: report of a WHO Expert Committee),127 pp.
American trypanosomiasis
Lent, H. & Wygodzinsky, P. Revision of the Triatominae (Hemiptera, Reduviidae) and their significance as vectors of Chagas disease. Bulletin of the American Museum of Natural History, 163: 125-520 (1979).
Miles, M.A. et al. Do radically dissimilar Trypanosoma cruzi strains (zymodemes) cause venezuelan and brazilian forms of Chagas disease? Lancet, i (June 20) 1338-1340 (1981).
PAHO. Status of Chagas disease in the Region of the Americas. Epidemiological Bulletin, 5, 5-9 (1984).
Zeledon, R. & Rabinovich, J.E. Chagas disease: an ecological appraisal with special emphasis on its insect vectors. Annual Review of Entomology, 26: 101-133 (1981).
A common and convenient way to group the leishmanial diseases of man, and their known or suspected vectors, is to divide them into those which produce mainly cutaneous lesions (including the clinically distinct - much more severe - mucocutaneous leishmaniasis) and those which cause a systemic infection with predominantly visceral lesions.
Visceral leishmaniasis in the Mediterranean basin and southwestern Asia, where infantile kala-azar mainly occurs, is caused by Leishmania (L.) infantum, of which there are many vectors; chiefly species of the subgenus Larroussius (Maps 39a, 42c, 42d, 43a).
In sub-Saharan Africa (Map 39a), kala-azar is caused by L.(L.) donovani complex organisms of more doubtful status, but which usually are grouped, together with the Indian parasites, as L.(L.) donovani. Human infections are rare, and the vectors unknown, in areas west of Sudan. To the South and East of Sudan, VL is occasionally reported in Somalia, Djibouti and Uganda. (Rare cases are reported as far south as Malawi, Zambia and Zaire, where nothing is known of either the vectors or parasites responsible). In eastern Africa, VL epidemics have occurred in Sudan, Kenya and neighbouring lowland areas of western Ethiopia, where another species of Larroussius, (P. orientalis) is the established vector. Species of the subgenus Synphlebotomus, (particularly P. martiniand P. celiae) are incriminated or probable vectors of L.(L.) donovani in Kenya, contiguous areas of Uganda and (probably) of Somalia and Ethiopia.
In the Indian subcontinent VL is caused by L.(L.) donovani only, with a single vector [P.(Euphlebotomus) argentipes], but in China both L.(L.) donovani and L.(L.) infantum occur, in different areas and with several different vectors (Maps 39b, 44). It has recently been demonstrated (isolates typed by monoclonar antibodies) that in northwest China (Turfan county) L.(L.) infantum is transmitted by P.(Pa) alexandri (Lihen, 1987).
Visceral leishmaniasis occurs over a very wide area in the Americas, in sporadic and often widely-separated foci, from Mexico to northern Argentina (Map 39c). Surprisingly, throughout this vast range there is only one apparent vector, Lutzomyia (Lutzomyia) longipalpis, often peridomestic in habits. The Neotropical visceral parasite is at present referred to as L.(L.) chagasi, but it undoubtedly includes parasites currently indistinguishable from L.(L.) infantum, accidentally imported from Europe in the past. The status of L.(L.) chagasi as an indigenous parasite of the New World is still uncertain and requires further study.
In the Old World anthroponotic cutaneous leishmaniasis (ACL) is caused by L.(L.) tropica (formerly L. tropica minor) and the distribution is shown in Map 40a. ACL is found in well-established settlements, often in urban conditions; there is no known silvatic reservoir and transmission is peridomestic. There is now convincing evidence that most or all of the human cutaneous leishmanial lesions found in the western Mediterranean area (southern France, Italy, Spain, Malta, Morocco and Algeria, shown as ACL in Map 40a) are cutaneous manifestations of infection with L.(L.) infantum, rather than with L.(L.) tropica as formerly was supposed. However, the causative agent of ACL in the eastern part of the Mediterranean and the western parts of Asia is L.(L.) tropica.
Of the zoonotic cutaneous leishmaniases (ZCL) of the Old World, that caused by L.(L.) major (formerly L. tropica major) is the most extensive and has the most known or suspected vectors (Map 40b). ZCL due to L.major is characteristic of lowland steppe and semi-desert zones. L.(L.) aethiopica, by contrast, is a silvatic species found only at high altitudes, around 2000m, mainly on the Ethiopian highland plateau, but also in Kenya (in the Aberdare range and on Mount Elgon). The normal hosts are various species of rock and tree hyrax. Occasional cases of cutaneous leishmaniasis occur in Tanzania, south of Lake Victoria, also in Zambia and Malawi; lesions resemble those seen in the Kenya and Ethiopian highlands, but nothing is known of the etiology, reservoir hosts or of possible vectors.
Two undescribed L. (Leishmania) spp. occur in hot, dry lowland areas of Namibia. One of these species is found in human lesions and in the sandfly P. (Synphlebotomus) rossi. The other parasite is found in a species of hyrax, also closely associated with P.rossi. The situation recalls that of L.(L.) aethiopica, since the natural hosts are also hyrax, with occasional human infections, but both of the organisms are biochemically distinct from L.(L.) aethiopica. Human CL of unknown etiology is quite common in western areas of Sudan and along part of the Nile north of Khartoum, but neither the parasites nor the possible vectors have been properly studied.
In the Neotropical region, the causative organisms of cutaneous leishmaniasis belong to either the L.(L.) mexicana complex, transmitted by Lutzomyia species of the subgenus Nyssomyia (Map 41b), or to the L.(V.) braziliensis complex, transmitted mainly by species of the subgenera Nyssomyia and Psychodopygus (Map 41a). The Nyssomyia species, which transmit L.(L.) mexicana complex parasites, are less anthropophilic than the sandflies (mainly species of Nyssomyia and Psychodopygus), which transmit the L.(V.) braziliensis complex parasites; hence, L. (V.) braziliensis complex parasites tend to be the more frequent cause of human CL infection.
Some members of, or allied to, the L. (L.) mexicana complex are associated with specific small foci (Map 41). These include L.(L.) venezuelensis (so far known from one small focus in the state of Lara, Venezuela); the vector is thought to be Lutzomyia (Nyssomyia) olmeca bicolor; L.(L.) gamhami, a parasite of uncertain affinity, occurs in the Venezuelan Andes at altitudes of 800 - 1800m, the likely vector being Lutzomyia townsendi; L.(L.) pifanoi, isolated from patients with diffuse cutaneous leishmaniasis in several areas of Venezuela, may include a number of different organisms. A parasite assigned to L.(L.) pifanoi was isolated from Lutzomyia (Nyssomyia) flaviscutellata; this particular parasite may well prove to be L.(L.) amazonensis, of which L.(N.) flaviscutellata is the established vector. A parasite also associated with diffuse cutaneous leishmaniasis in the Dominican Republic has not been named, but appears to belong in the mexicana complex.
The phlebotomine sandflies (Diptera: Family Psychodidae, sub-family Phlebotominae) are the vectors of the various leishmanial diseases of man and other mammals. Of some 700 phlebotomine species and subspecies, about 70 are considered to be vectors of Leishmania parasites to man. All known or suspected vectors belong either to the genus Phlebotomus (with about 90 species in the Old World: Africa, Asia, Europe, Australasia), or to the genus Lutzomyia (with about 300 New World species, in the Americas). A list of vectors by country is to be found in a WHO Expert Committee report on the leishmaniases (WHO 1984).
The main North African vectors of visceral leishmaniasis are P. (Larroussius) pemiciosus and P. (Larr.) longicuspis, but P. (Larr.) lanqeroni also occurs, from Morocco eastward to Egypt, and is almost certainly a vector of VL near Alexandria (Map 39a). Other Larroussius vectors, in the eastern Mediterranean area, are P. (Larr.) syriacus [= P. major syriacus] and P.(Larr.) tobbi (Maps 42c and 43a); P. (Larr.) smirnovi is considered a VL vector in the southern USSR, together with P. (Adlerius) longiductus (Map 43c).
Recent information from China reveals that the extent and complexity of the visceral leishmaniasis and vector situation in that vast country is greater than was apparent when Map 39b was prepared. The situation as now known is shown in Map 44.
In the low-lying and densely populated North China Plain (mainly below 200m altitude), anthroponotic L.(L.) donovani is found, with P. (Adlerius) chinensis as the known vector. Smaller discrete foci of anthroponotic L. donovani and P. chinensis lie further south, between the Chinling mountains and the Yangtze river.
To the west and north of the North China Plain, on the higher loess plateau (altitude about 500m, L.(L.) infantum occurs, with P. chinensis as the vector; both domestic dogs and a silvatic carnivore, the racoon dog (Nyctereutes procyonoides), are reservoir hosts in this extensive region.
North and west of the line on Map 44 which marks the known northern and western limits of P. chinensis, close to the western end of the Great Wall, are several scattered VL foci where the parasite is shown as L. (L.) infantum, with Nyctereutes procyonoides as a silvatic reservoir host and, it is believed, P. (Larroussius) wui [= P. major wui] is the vector. Further east, north of Beijing, VL parasites have been isolated from the racoon dog also, but the vector in this area seems more likely to P. chinensis sensu lato.
Human VL is found also in the higher areas of Szechuan Province, southwest of the loess plateau; it is unclear whether the parasite in this region is L. donovani or L.infantum, but the vector here is also P. (Adlerius) chinensis, except that this species is replaced, at altitudes over 900m in the Szechuan mountains, by the closely related P. (Adlerius) sichuanensis. This sandfly is found up to altitudes of 2800m in Szechuan: human VL transmission continues to over 2000m.
In the more arid foothill areas of northwestern China, in the Autonomous Region of Sinkiang (Xinjiang), which lies north of the Tibetan Plateau and includes the Tarim and Dzungaria basins (separated by the Tien Shan mountains which run west east between them), are several foci of what may be anthroponotic L.(L.) donovani. In one of these foci at least, P. (Paraphlebotomus) alexandri was demonstrated to be the principal vector to man, but in different altitudinal and ecological zones of the same area, both P. (Adlerius) longiductus and P. (Larroussius) wui are also involved in the natural history of the infection. The extensive distribution of P. (Paraphlebotomus) alexandri (not shown on the maps for reasons of space and clarity) stretches from North Africa and the western Mediterranean eastward to central China.
The most important visceral leishmaniasis vector of southern Asia is currently P. argentipes, in the Indian subcontinent. This is the only member of the subgenus Euphlebotomus so far implicated in transmission of human disease, although only in that part of its extensive
range (Maps 39b, 43d). In terms of numbers of VL cases, P. argentipes is undoubtedly now the most important sandfly vector of all. P. argentipes is possibly a species-complex; as at present defined the species is geographically variable, in both morphology and biology, over its extensive range; the man-biting Indian vector form can be distinguished from the non-anthropophilic populations of South-East Asia (where leishmaniasis is unknown).
One species of the subgenus Synphlebotomus, (P. martini), is the most widespread vector in Kenya, contiguous areas of Uganda and probably of Somalia and Ethiopia. Two sibling species of P. mantini (P. celiae and P. vansomerenae) may act as additional vectors in limited areas of eastern Kenya, but particularly in southern Ethiopia and southern Somalia, where P. celiae usually occurs together with P. martini, but P. vansomerenae is absent. (Maps 39a, 43b).
In the Old World, P. sergenti and P. papatasi are the main vectors of anthroponotic cutaneous leishmaniasis (due to L.(L.) tropica), but P. perfiliewi is considered to be the vector in Italy and is probably important in neighbouring countries (Maps 40a, 42a, 42b, 42d).
The most important and widespread vector of zoonotic cutaneous leishmaniasis (ZCL) caused by L.(L.) major is P. (Phlebotomus) papatasi; other species are mainly of regional or local importance, such as P.(Phlebotomus) salehi, or principally transmit the parasite between its silvatic rodent hosts, but occasionally also to man, such as P.(Phlebotomus) duboscqi (Maps 40b, 42a). In Ethiopia the main vector of L.(L.) aethiopica is P.(Larroussius) longipes (Map 42c), but the closely related P. pedifer becomes important in the southern part of the Ethiopian highlands and is the main vector in the Kenya foci (Maps 40b, 43a).
In the Neotropical region, parasites of the Leishmania (L.) mexicana complex are transmitted by Lutzomyia species of the subgenus Nyssomyia, such as L.(N.) flaviscutellata and L.(N.) olmeca olmeca (Map 41b); other members of the same subgenus are among the many vectors of the distinct Leishmania (V.) braziliensis complex of species (these include Lutzomyia (N.) umbratilis, the vector of Leishmania (V.) guyanensis, and L. (N.) trapidoi and L. (N.) ylephiletor, vectors of Leishmania (V.) panamensis: Map 41a). The vectors of the braziliensis complex of parasites are taxonomically diverse, but species of the subgenera Nyssomyia (see above) and Psychodopygus (e.g. L. (Psychodopygus) wellcomei, a vector of L.(V.) braziliensis), are among the most important. L. cruciata and L. gomezi are cutaneous leishmaniasis vectors which belong to the cruciata group, not allocated to any subgenus (Map 41a). L. diabolica (distribution not shown) is also a member of the cruciata group; it occurs in Mexico and the southern USA; this species is suspected to be the vector of the few cases of human CL recorded from Texas (parasites all of L.(L.) mexicana type). The vectors of L.(V.) peruviana are not known with certainty, but prime suspects include one or more of three species: L.(Helocyrtomyia) peruensis, L. verrucarum, a member of the verrucarum group of species, (see Map 41a) and L.(Helocyrtomyia) noguchii (not shown in Map 41a).
Maps 42 and 43, which show in greater detail the distribution of some Old World vectors by subgeneric groups, rely heavily on the compilations of Lewis (1982) with some later additions. The distribution of leishmaniasis vectors in the Americas is not known in sufficient detail (for many species) to produce similar maps, but Martins and co-workers (1978) mapped the distribution, as then known, for a number of important species. Young and Perkins (1984) mapped the phlebotomines of continental America north of Mexico.
1 The maps which accompany this text were prepared some time before the recent revision of the genus Leishmania by Lainson and Shaw (1987). For technical reasons, it was not possible to change the names on the body of the maps, which use the subspecific names and terminology valid before the recent review. The text, however, uses the new (1987) subgeneric classification, in which former subspecies are given specific rank and the former Section Peripylaria (which included the subspecies of the L. braziliensis complex) is now raised to a subgenus, Viannia. Old and new names are tabulated in the Appendix). [NOTE: The reptilian parasites of the former Section Peripylaria are no longer considered to be Leishmania].
INDEX OF MAPS
The maps listed below are presented on pages 101 - 106.
MAP 39a. Visceral leishmaniasis in the Mediterranean basin, East and West Africa, and southwestern Asia, and vectors.
MAP 39b Visceral leishmaniasis in the Indian subcontinent and China, and vectors.
MAP 39c Visceral leishmaniasis in the Americas, and vectors.
MAP 40a Anthroponotic cutaneous leishmaniasis and vectors in the Old World.
MAP 40b Zoonotic cutaneous leishmaniasis and vectors in the Old World.
MAP 41a Distribution and vectors of cutaneous leishmaniasis in the Neotropical region (L. (V.) braziliensis complex) .
MAP 41b Distribution and vectors of L.(L.) mexicana parasite complex, including the Venezuelan foci.
MAP 42-43 Distribution of Old World vectors/leishmaniasis.
MAP 44 Visceral leishmaniasis and vectors in China.
List of phlebotomine subgeneric abbreviations used in the map legends:
Lainson, R. The American leishmaniases: some observations on their ecology and epidemiology. Transactions of the Royal Society of Tropical Medicine and Hygiene, 77: 569-596 (1983).
Lane, R.P. Phlebotomine sandflies. In: Manson's tropical diseases, 19th edition, Eds. P.E.C. Manson-Bahr & D.R. Bell, London, Bailliere Tindall,1987, pp.1395-1404.
Lewis, D.J. The phlebotomine sandflies (Diptera: Psychodidae) of the Oriental Region. Bulletin of the British Museum (Natural History), Entomology Series, 37: 217-343 (1978).
Lewis, D.J. A taxonomic review of the genus Phlebotomus (Diptera: Psychodidae). Bulletin of the British Museum (Natural History, Entomology Series, 45: 121-209 (1982).
Lewis, D.J. & Ward, R.D. Transmission and vectors. In: The leishmaniases in biology and medicine [vols.1 & 2]. Eds. W. Peters & R. Killick-Kendrick, London & New York, Academic Press,1987, pp. 235-262.
Li-hen, G., Yong-Xiang, X., Bao-shan, L. & Jiang, D. The role of Phlebotomus alexandri Santon, 1928, in the transmission of kala-azar. Bulletin of the World Health Organization, 64: 107-112 (1986).
Martins, A.V., Williams, P. & Falcao, A.L. American sandflies (Diptera: Psychodidae, Phlebotominae). Rio de Janeiro, Academia Brasileira de Ciencias, 1978, 195 pp.
Minter, D.M. The Trypanosomatid flagellates: genera Trypanosoma and Leishmania. In: Manson's tropical diseases, 19th edition, Eds. P.E.C. Manson-Bahr & D.R. Bell, London, Bailliere Tindall,1987, pp. 1256-1322.
WHO. Studies on leishmaniasis vectors/reservoirs and their control in the Old World; Parts I to V. (Compiled by A.R. Zahar). WHO, Geneva, unpublished documents: Parts I & II (1979). General Review, Europe and North Africa. WHO/VBC/79.749. 88 pp. + corr. 1 & 2; Part III (1980a). Middle East. WHO/VBC/80.776, ii + 78 pp. Part IV (1980b). Asia & Pacific. WHO/VBC/80.786, 85 pp. Part V (1981). Tropical Africa. WHO/VBC/81.825, 198 pp.
WHO. Epidemiology of the leishmaniases. Report of the Third Meeting of the Scientific Working Group on Leishmaniases. Unpublished WHO document, TDR/Leish-SWG(3)/81.3, 41 pp. (1981).
WHO. Sixth Annual Report of the Special Programme for Research and Training in Tropical Diseases. Chapter 7: Leishmaniases. Unpublished WHO document TDR/PR-6l837-LEISH, pp 193-205 (1983).
WHO Technical Report Series, No. 701,1984 (The leishmaniases, report of a WHO Expert Committee), 140 pp.
Young, D.G. & Perkins, P.V. Phlebotomine sand flies of North America (Diptera: Psychodidae). Mosquito News, 44: 263-304 (1984).
The causative agent is the bacterium, Yersinia pestis. Plague persists in rodent populations in many parts of the world in an enzootic (asymptomatic) form. Occasionally, the disease is transmitted by fleas from these enzootic hosts to several species of plague-susceptible squirrels, chipmunks, wood rats, marmots and prairie dogs, which may then suffer epizootic plague and die in vast numbers.
Urban rat-borne plague remains a potential threat wherever commensal rat species come into contact with either the enzootic rodent species or the epizootic plague-susceptible rodent species in urban or peri-urban areas. The transfer of Y. pestis organisms from the native rodent to the commensal rats by exchange of fleas occurs rather easily. Once commensal rodents become infected, the risk of human populace is greatly increased.
Human plague within the last decade has been reported from 10 countries in Africa: Angola, Kenya, Libyan Arab Jamahiriya, Madagascar, Mozambique, South Africa, United Republic of Tanzania, Uganda, Zaire and Zimbabwe; five in the Americas: Bolivia, Brazil, Ecuador, Peru and the USA; and three in Asia: Burma, China and Viet Nam (WHO,1987). The areas where plague may occur are shown in Map 45. In 1986, 1003 cases of human plague including 115 deaths, recorded from 8 countries of the world, were notified to WHO (WHO, 1987). Nine countries had reported a total of 483 cases including 51 deaths in 1985. During 1986, 700 cases of human plague, nearly 70% of the world total, occurred in 2 African countries, Uganda and the United Republic of Tanzania.
Plague is one of the diseases listed under the International Health Regulations of the World Health Organization, which require that ships be periodically "deratted" and rat-proofed to stop the spread to plague-free areas.
In plague transmission, fleas subsisting on infected animals and frequenting their nests and burrows become "blocked" by the bacilli multiplying in the ingested blood-meal and forming a viscous accumulation which plugs the proven triculus. Thereafter, when the flea makes attempts to feed, the oesophageal contents contaminated with bacilli are regurgitated into the new victim of the abortive flea-bite. During the 2-10 days before a blocked flea dies, it can infect dozens of animals, or men. Effective vector species are those in which blood-meals frequently cause blockage.
Fleas capable of biting man and potentially infected rodents may be regarded as possible vectors, but flea species vary very much in their ability to transmit the disease.
Xenopsylla cheopis, the oriental rat flea is the most important vector of urban plague and murine typhus. Having probably originated in Egypt, it has been distributed to all parts of the world in ships' cargoes, particularly in the second half of the 19th century. The spread inland from the ports of entry continues and now extends to northern USA, the ports of Gdynia (Poland) and Batum (USSR), and Manchuria (China) and the Republic of Korea in northeastern Asia (Map 46a). It is now characteristically found in buildings parasitizing R. rattus, R. norvegicus and other species of Rattus .
Xenopsylla astia, a parasite of gerbils and rats, ranges from Arabia and Iran through India to South-East Asia (Map. 46b). Less domestic and a weaker plague vector than X. cheopis, it has been introduced on the east coast of Africa.
Xenopsylla brasiliensis is native to all Africa, south of the Sahara, but it has spread to other parts of the world, notably Brazil and India (Map 46b). It is an effective plague vector, especially in rural environments.
Nosopsyllus fasciatus, the northern rat flea, is prevalent on rats in Europe, temperate North America and Australia. It is a relatively unimportant vector of plague.
Pulex irritans, the human flea, is an Old World species that has become virtually ubiquitous (Map 46c). Its range of hosts is wide. Piggeries, after the pigs have been taken to market, are often the source of acute human infections. Laboratory tests indicate that it has all the qualities of a plague vector, and in certain areas of human plague (e.g. Kurdistan), it was the sole and abundant domestic flea.
The rickettsiae ingested from the rats proliferate in the flea's stomach and malpighian tubules, and are then excreted along with its faeces for the rest of its life. Human infection results from contamination from the dried faeces and crushed bodies of the fleas.
The vectors are the same as those of plague described above.
INDEX OF MAPS
The maps listed below are presented on pages 111 - 113.
MAP 45 Known and probable foci of plague transmission to man.
MAP 46 Distribution of plague vectors Xenopsylla cheopis (46a); X. astia and X. brasiliensis (46b); Pulex irritans (46c).
MAP 47 Flea-borne (Murine) typhus (Rickettsia mooseri) endemic areas.
REFERENCES
Alexander, J.O. Arthropods and human skin. Springer-Verlag, Berlin, 1984.
Gratz, N.G. & Brown, A.W.A. Fleas - biology and control. WHO unpublished document, WHO/VBC/84.874, 1983, 46 pp.
Holland, G.P. Evolution, classification and host relationships of Siphonaptera. Annual Review of Entomology, 9: 123-146 (1964).
Pollitzer, R. Plague. Geneva, World Health Organization, Monograph Series, No. 22, 1954.
Traub, R. & Starke, H. (ed.) Fleas: Proceedings of the International Conference on Fleas, Peterborough,1977. Rotterdam, Balkema,1980.
WHO. Human plague in1986. Weekly Epidemiological Record,62: 299-300 (1987).
Transmission of O. volvulus is due entirely to diurnally active female Simulium blackflies of several species groups. Simulium always breed in running water so that the distribution of onchocerciasis is generally associated with rivers and streams. The larvae and pupae congregate on substrates where the water is most highly oxygenated, so that breeding sites are concentrated at rapids, waterfalls and spillways of dams. Since female Similium are strong fliers, and many people make daily visits to watercourses, onchocerciasis may be endemic for more than 20 km away from the vector breeding sites. Infective females of S. damnosum (sensu lato) have been found to disperse over hundreds of kilometres in West African savannas, giving rise to long range transmission and serious reinvasion of areas covered by vector control activities of the WHO Onchocerciasis Control Programme (OCP) involving initially seven countries and now extended to a further four countries (WHO, 1981). The OCP is based on larvicidal control of onchocerciasis vectors breeding in over 24 000 km of infested rivers (WHO, 1985).
Map 49 shows the overall distribution of onchocerciasis in Africa and the Yemen where transmission is due to two main groups of vectors: the Simulium (Edwardsellum) damnosum group (Map 50) and the Simulium (Lewisellum) neavei group (Map 51). The vectors of the former group are much more widespread and important, with a great complexity of sibling species.
The S. neavei group comprises about a dozen species which breed in streams of hilly and mountainous areas around the Zaire basin and in eastern Africa. The larvae and pupae invariably attach themselves phoretically to freshwater crabs of particular species. The S. neavei group includes three distinct vectors of upland onchocerciasis (Map 51): S. ethiopiense in the southern highlands of Ethiopia: S. woodi in the Tanzanian highlands and S. neavei (sensu stricto) in Uganda and around the Zaire basin and formerly in Kenya, where eradication of both vector and disease was accomplished during the 1950s (Raybould & White, 1979).
The S. damnosum group comprises about 30 known sibling species in Africa. Species identification depends largely on cytotaxonomic characteristics of salivary polytene chromosomes (Quillevere, 1979). Most of these species are not anthropophilic and are little known. The following seven members of the S. damnosum group are regarded as important vectors of African onchocerciasis.
S. damnosum (sensu stricto) and S. sirbanum breeding in large rivers of Uganda, southern Sudan and across the West African savannas (Maps 52a, 52b); responsible for transmission of the most pathogenic form of O. volvulus;
S. squamosum and S. yahense breeding in small forested rivers of West-Africa (Maps 52c, 52d) and responsible for transmission of less pathogenic O. volvulus;
S. sanctipauli and S. soubrense breeding in large rivers of both forest and savanna zones in West Africa; responsible for transmission of both mild and severe forms of O. volvulus;
S. kilibanum breeding in mountain streams of central and eastern Africa, in Uganda, on the Burundi/Zaire border and probably in Tanzania is responsible for transmission of a less pathogenic form of the parasite in cool highland situations.
The only other African vector of human onchocerciasis is S. albivirgulatum which breeds in slow-flowing rivers in forested areas of several countries and is apparently responsible for transmission of O. volvulus in parts of central Zaire where the disease is endemic, but the S. neavei group and the S. damnosum group are absent (Fain et al.,1969).
Scattered foci of human onchocerciasis in tropical American countries have various vectors which are not well understood ecologically or taxonomically. Presumably these vectors are as complex as the S. damnosum group in Africa, but cytotaxonomic analysis is only now beginning. Moreover, new foci of onchocerciasis are still being found in the Americas. There are clinical differences in the disease in Middle and South America (Suzuki,1983; Tada,1983). The principal known vectors are shown on Map 53.
S. metallicum resembles S. damnosum morphologically (although it is not closely related taxonomically) and breeds in medium to large rivers (Map 53a). It transmits 0. volvulus in several foci in Venezuela. Different sibling species are likely in these localities, since the biology of S. metallicum is variable and it occurs extensively where onchocerciasis is absent. Two have been named S. jobinsi and S. pingi.
The S. amazonicum group comprises at least six species in tropical America, only one of which - S. oyapockense - is anthropophilic. It transmits 0. volvulus in areas around the border between southern Venezuela and northern Brazil (Map 53b). Breeding of S. oyapockense occurs in large forested rivers, but the immature stages are elusive.
S. exiguum is a vector of a variable importance, being secondary to S. metallicum in Venezuelan foci, but the sole vector in foci in southern Colombia and an important vector in northern Ecuador (Map 53c). This blackfly is widespread in onchocerciasis-free areas and it can be assumed that S. exiguum is a species complex with only one or two vectors among the sibling species. Breeding sites of S. exiguum are also variable .
S. ochraceum and its two taxa breed in very small streams and seepages, the larvae hiding under fallen leaves and herbage. It is the principal vector of 0. volvulus in Mexico and Guatemala (Map 53c), feeding predominantly on the head, neck and shoulders, so that onchocercal nodules tend to occur on upper parts of the patient's body. Due to the scattered obscure breeding sites of S. ochraceum, control by means of larvicides is highly labour intensive.
Secondary vectors of 0. volvulus in some foci are S. metallicum and possibly S. callidum and S. gonzales in Central American countries and S. quadriviffatum in South American countries (Map 53b).
INDEX OF MAPS
The maps listed below are presented on pages 118 - 126.
MAP 48 Geographical distribution of onchocerciasis in Latin America.
MAP 49 Geographical distribution of onchocerclasis in Africa and the Arabian Peninsula.
MAP 50 Approximate known distribution of Simulium damnosum s.l.
MAP 51 Geographical distribution of the Simulium neavei group.
MAP 52a Pre-control range of S. damnosum (sensu stricto) in West Africa.
MAP 52b Pre-control range of S. sirbanum distribution in West Africa.
MAP 52c Pre-control range of S. squamosum in West Africa.
MAP 52d Pre-control range of S. yahense in West Africa.
MAP 53a Known foci of S. metallicum and S. guianense in Latin America.
MAP 53b Known foci of S. oyapockense and S. quadrivittatum in Latin America and the Caribbean islands.
MAP 53c Known distribution of S. exigeum and S. ochraceum in Latin America and the Caribbean islands.
REFERENCES
Crosskey, R.W. The vectors of human onchocerciasis and their biology. In: Mackenzie, C.D. & Nelson, G.S. eds. Human onchocerciasis. London, Academic Press,1989.
Fain, A, Wery, M. Tilkin, J. Recherches sur les filarioses humaines dans la region de la Cuvette Centrale (Republique democratique du Congo). Annales de la Societe belge de Medecine tropicale, 49: 629648(1969).
Quillevere, D. Contribution l'etude des caracteristiques taxonomiques, bioecologiques et vectrices des membres du complexe Simulium damnosum presents en Cote d'Ivoire, Paris, ORSTOM,1979, 304pp.
Raybould, J.N. & White, G.B. The distribution, bionomics and control of onchocerciasis vectors (Diptera: Simuliidae) in Eastern Africa and the Yemen. Tropenmedizin und Parasitologie, 30: 505-547,1979.
Suzuki, T. A guidebook for Guatemala on onchocerciasis (Robles' disease). Guatemala, Guatemalo-Japan Cooperative Project on Onchocerciasis Research and Control, 1983, 155 pp.
Tada, I. ed. A comparative study on Onchocerciasis between South and Central Americas. Kumamato, Shimoda, 1983, 79 pp.
WHO. Senegambia Project: Onchocerciasis control in Guinea, Guinea-Bissau, Mali, Senegal and Sierra Leone, Geneva, unpublished WHO document VBC/81.2; AFR/lCP/MPD/007, 1981, 122 pp.
WHO. Ten years of onchocerciasis control in West Africa: Review of the Work of the Onchocerciasis Control Programme in the Volta River Basin area from 1974 to 1984, Geneva, unpublished WHO document OCP/GVA/85.1B, 1985, 113 pp.
WHO. Technical Report Series No.752, 1987. (Third report of the WHO Expert Committee on Onchocerciasis). Geneva, World Health Organization, 167 pp.
Its clinical manifestations are often delayed for months or years and can continue for up to 17 years. They are caused by migration of the adult worm through the subcutaneous and deeper tissues of the body, leading to recurring or Calabar swellings and the worms may migrate under the bulbar conjunctivae. Allergic reactions, arthritis and neurological symptoms can occur. Diagnosis may be complicated by the presence of other filarial infections, especially onchocerciasis (Fain, 1981). The microfilarial have a day-time periodicity.
A simian species of Loa loa with a nocturnal periodicity occurs in Mandrillus and other monkeys and may be the cause of the so-called Kampala eye worm in man in Uganda and also in Ethiopia.
Chrysops is a genus of the Tabanidae and over 40 species have been identified in Africa alone (Crosskey, 1980). The genus breeds mainly in forest swampland where the eggs are laid above water, into which the larvae drop. The life cycle requires four months or more to complete. Only the female fly bites man: the male Chrysops does not suck blood.
The principal vector species are Chrysops silacea and C. dimidiata, and the secondary vectors C. distinctipennis which has a widespread distribution and C. zahrai limited to southern Nigeria and Cameroon. These are all daytime feeders (Duke, 1955). Simian loiasis may be carried to man by C. streptobaluis in Ethiopia and C. Iongicornis in the Sudan. Other vectors of simian loiasis are C. Iangi in West Africa and C. griseicollis in West and Central Africa.
INDEX OF MAPS
The map listed below is presented on page 129.
Map 54 The distribution of loiasis and its vectors.
REFERENCES
Crosskey, R. ed. Catalogue of the Diptera of the Afrotropical Region, London, British Museum (Natural History), 1980, pp 281 - 283.
Duke, B.O.L. IV The development of Loa in flies of the genus Chrysops and the probable significance of the different species in the transmission of loiasis. Transactions of the Royal Society of Tropical Medicine and Hygiene, 49: 115-121 (1955).
Fain, A. Epidemiologie et pathologie de la loase. Annales de la Societe belge de Medecine tropicale, 61. 277-285 (1981).
Rhodain, F. Hypotheses concernant l'ecologie dynamique des infections a Loa. Bulletin de la Societe de Pathologie exotique et de ses filiales, 73: 182-191 (1980).
ACKNOWLEDGEMENTS
Grateful acknowledgement is made to the following persons, in addition to the contributors who reviewed the manuscript and made comments, or otherwise collaborated in the preparation of this document:
Dr. P.F. Beales, Chief, Programme and Training, Malaria Action Programme, World Health Organization, Geneva, Switzerland; Dr. J. Bektimirov, Assistant Director-General, World Health Organization, Geneva, Switzerland; Mr. R. Bos, Division of Vector Biology and Control, World Health Organization, Geneva, Switzerland; Professor A.N. Clements, Director of the Department of Entomology, London School of Hygiene and Tropical Medicine, London, England; Dr. R.W. Crosskey, British Museum (Natural History), London, England; Professor G. Davidson (retired), formerly Head of the Department of Entomology, London School of Hygiene and Tropical Medicine, London, England; Dr. A. Davis, Director, Parasitic Diseases Programme, World Health Organization, Geneva, Switzerland; Dr. P. Desjeux, Trypanosomiases and Leishmaniases, World Health Organization, Geneva, Switzerland; Dr. J. Esparza, Biomedical Research, Global Programme on AIDS, World Health Organization, Geneva, Switzerland; Professor A. Farhang Azad, Department of Microbiology and Immunology, School of Medicine, University of Maryland, United States of America; Dr. N.G. Gratz (retired), formerly Director, Division of Vector Biology and Control, World Health Organization, Geneva, Switzerland; Dr. I. Guendon, Microbiology and Immunology Support Services, Division of Communicable Diseases, World Health Organization, Geneva, Switzerland; Dr. J. Haworth (retired), formerly Chief, Publications, World Health Organization, Geneva, Switzerland (Editor); the late Harry Hoogstraal, formerly United States Naval Medical Research Unit No. 3, Cairo, Egypt; Dr. I.A.H. Ismail, Delivery and Management of Vector Control, World Health Organization, Geneva, Switzerland (Operational Officer Responsible); Dr A.B. Knudsen, Chief, Delivery and Management of Vector Control, World Health Organization, Geneva, Switzerland; Mr. F.A.S. Kuzoe, Trypanosomiases and Leishmaniases, World Health Organization, Geneva, Switzerland; Dr. C.J. Leake, Department of Entomology, London School of Hygiene and Tropical Medicine, London, England; Dr. R. Le Berre, Research Capability Strengthening, Special Programme for Research and Training in Tropical Diseases, World Health Organization, Geneva, Switzerland; Dr. J. McDade, Director of the WHO Collaborating Centre on Rickettsial Diseases, Centers for Disease Control, Atlanta, Georgia, United States of America; Dr. J.D.M. Marr, Onchocerciasis Control Programme in West Africa, World Health Organization, Geneva, Switzerland; Dr. F. Modabber, Trypanosomiases and Leishmaniases, World Health Organization, Geneva, Switzerland; Dr. A. Moncayo, Trypanosomiases and Leishmaniases, World Health Organization, Geneva, Switzerland; Dr. D.A. Muir, Malaria Action Programme, World Health Organization, Geneva, Switzerland; Dr. J.A. Najera-Morrondo, Director, Malaria Action Programme, World Health Organization, Geneva, Switzerland; the late Dr. C.P. Pant, formerly Chief, Ecology and Control of Vectors, World Health Organization, Geneva, Switzerland; Dr. Y. Pervikov, Microbiology and Immunology Support Services, Division of Communicable Diseases, World Health Organization, Geneva, Switzerland; Dr. P. de Raadt, Chief, Trypanosomiases and Leishmaniases, World Health Organization, Geneva, Switzerland; Dr. P. Ranque, Chief, Filarial Infections, World Health Organization, Geneva, Switzerland; Dr. N. Rishikesh, Chief, Development of Vector Control Technology, World Health Organization, Geneva, Switzerland; Dr. A. Smith (retired), formerly Division of Vector Biology and Control, World Health Organization, Geneva, Switzerland; Dr. E.D. Tikhomirov, Microbiology and Immunology Support Services, Division of Communicable Diseases, World Health Organization, Geneva, Switzerland; Dr. R.J. Tonn (retired), formerly Chief, Planning, Management and Operations, Division of Vector Biology and Control, World Health Organization, Geneva, Switzerland; Mrs S. Walton, Division of Vector Biology and Control, World Health Organization, Geneva, Switzerland (secretarial assistance); Dr. D. Woodall, Development of Epidemiological and Health Statistical Services, World Health Organization, Geneva, Switzerland; and Dr. A.R. Zahar (retired), formerly Division of Malaria Eradication and other Parasitic Diseases, World Health Organization, Geneva, Switzerland.
3. MOSQUITO BORNE VIRUS DISEASES
3.1 Yellow Fever (YF) (Urban yellow fever, silvatic or jungle yellow fever)
M. G. R. Varma
Pathogen: Yellow fever virus, Family Flaviviridae (previously
Togaviridae).
Vectors: Urban yellow fever, Aedes aegypti (Map 14);
jungle/rural yellow fever in Africa, Aedes africanus (Map 15b), A.
luteocephalus (Map 15c), A. simpsoni complex (Map 15d), A.
furcifer (Map 15e), A. taylori (Map 15f). Jungle yellow fever
in Central and South America, Haemagogus leucocelaenus (Map 16a),
H. janthinomys (Map 16b), H. spegazzinii (Map 16c) and
Sabethes chloropterus (Map 16d).
Reservoirs: Old World monkeys of the Family Cercopithecidae and
Colobidae;the bush-baby Galago senegalensis. In the New World
monkeys of the family Cebidae, especially species of Alouatta
(howler monkeys), Ateles (spider monkeys), Cebus, Aotes
and Callithrix. The basic reservoir is now considered to be the
mosquito vector.
Yellow fever is endemic in tropical Africa between 1 5\'a1N and 1 5\'a1S and in the north and eastern parts of South America and epidemics occur in parts of Middle America (Map 13c). As did the main vector, A. aegypti, the disease originated in Africa and was carried along with the vector to the New World with the slave trade in the 16th century. The African slaves did not appear to suffer severely from yellow fever but mortality among the Europeans was high. Serious epidemics occurred in the coastal towns of South America and the USA as far north as New York. More recently, there was a spectacular outbreak in Ethiopia in 1960-1962 with at least an estimated 15 000 deaths, and in 1986/87 severe outbreaks occurred in West Africa. All the urban epidemics in the New World were transmitted by A. aegypti and were severe, but at some time, the virus "escaped" into the forests and in 1932 the disease was discovered in the interior of Brazil from where A. aegypti had been eradicated. Investigations led to the recognition of jungle yellow fever maintained by forest-canopy inhabiting monkeys and mosquitos which breed in the canopy. Urban yellow fever has now been controlled by immunization, but little progress has been made in reducing the risk of rural yellow fever and in the enzootic areas of Africa and America, human cases occur every year. The urban vector, A. aegypti is re-invading areas in South America from where it had been eradicated.3.2 Dengue and Dengue Haemorrhagic Fever (DHF) (Dengue shock syndrome, DSS, Break-bone fever)
M. G. R. Varma
Pathogen: Arbovirus, Family Flaviviridae (previously Togaviridae);
four serotypes, dengue 1, 2, 3 and 4.
Vectors: Mosquitos, particularly Aedes aegypti (Map 14),
A. albopictus, (Map 15a), A. scutellaris (Map 11) and A.
polynesiensis (Map 11).
Reservoirs: Not defined but probably man and possibly forest
monkeys in Malaysia.
Dengue is endemic throughout the tropics, particularly Asia, the Pacific and Caribbean (Map 13 a, b) but tends to occur in periodic epidemics. In its classic form, it is a benign disease characterized by biphasic fever, myalgia or arthralgia and rash.3.3 Japanese Encephalitis (JE) (Japanese B encephalitis, Russian autumnal encephalitis, summer encephalitis)
M. G. R. Varma
Pathogen: Japanese encephalitis virus, Family Flaviviridae
(previously Togaviridae).
Vectors: Mosquitos, principally Culex tritaeniorhynchus, C.
gelidus and C. vishnui group.
Reservoirs: Probably birds, but night herons, egrets and
domestic pigs also act as amplifying hosts of the virus.
Japanese encephalitis has caused widespread epidemics in Japan and the Republic of Korea. The known distribution of the disease extends from maritime Siberia, through Japan, Republic of Korea, China, Indonesia, Singapore, Malaysia, Thailand, Viet Nam, Burma, Nepal, India and Sri Lanka to the west coast of India (Map 17a). Recent epidemics have occurred in India since 1973; one in 1977-1978 in particular was characterized by high mortality.3.4 Other encephalitides
G. B. White3.4.1 California Encephalitis (CE) Group (Family Bunyaviridae)
Vectors: Aedes, Anopheles, Culex, Culiseta and Psorophora
spp. of mosquitos.
Reservoirs: Rodents and lagomorphs.
About 14 viruses are known in this Group, the name being misleading because human cases are rare in California. Apart from the Americas, cases of the disease have been reported from Africa and Europe and there has been a recent isolate of Snow Shoe Hare-like virus in China. Generally they cause sub-clinical symptoms in man, but occasional acute cases occur. Rodents and lagomorphs are the main wild hosts and reservoirs in North America, where ten types of virus in this Group occur.3.4.2 Eastern Equine Encephalitis (EEE) (Family Flaviviridae, genus
Vectors: To man chiefly Aedes (Ochlerotatus)
Reservoirs: Birds, possibly horses.
EEE is widespread in the Americas (Map 18d) with outbreaks occurring in eastern coastal areas of Canada, USA, through the Caribbean and South America as far south as Argentina. Horses are the main victims and human cases are sporadic, normally not exceeding 100 in any outbreak. The virus is enzootic in birds (introduced game-birds such as pheasants being especially susceptible) and has been isolated repeatedly in the Americas. Transmission among birds is largely due to ornithophilic Culex spp. and Culiseta spp. in which transovarial transmission also occurs. Vectors to man and horses are chiefly species of Aedes (Ochlerotatus).3.4.3 St. Louis Encephalitis (SLE) (Family Flaviviridae, genus Flavivirus)
Vectors: Culex spp.
Reservoirs: Birds.
SLE is widespread in the Americas (Map 18b) but human cases are rare though outbreaks may involve a number of patients. Symptoms are milder than for EEE or VEE and most infections are inapparent. Transmission is mainly due to Culex tarsalis (Map 19d) (western USA), C. quinquefasciatus (mid USA) and C. nigripalpus (Map 20c) (USA). Transovarial transmission is the main method of overwintering for the virus, which amplifies in birds.3.4.4 Western Equine Encephalitis (WEE) (Family Flaviviridae, genus Alphavirus)
Vectors: Culex spp., Culiseta spp.
Reservoirs: Passerine birds; possibly, reptiles and amphibia.
WEE is also widespread in Middle and South America and isolates have been found in Europe, but serious outbreaks are confined to North America and no human cases have been detected in Europe (Map 18c). WEE virus has been isolated in Culex, Aedes, Anopheles, Culiseta and Psorophora, but transmission is mainly due to Culex tarsalis (Map 19d) in western USA. WEE is important as a horse disease; outbreaks usually involve no more than a few hundred human cases with moderate symptoms and the majority of infections are inapparent.3.4.5 Venezuelan Equine Encephalitis (VEE) (Family Flaviviridae, genus Alphavirus)
Vectors: Aedes spp., Culex portesi, Psorophora ferox
Reservoirs: Rodents.
VEE resembles WEE in many respects, except that the reservoir is in rodents. Many species of mosquitos are probably involved during epidemics, the virus having been isolated from 26 species. In North America, important vectors are breeding in salt marshes: Aedes taeniorhynchus (Map 20a), A. sollicitans (Map 20b), Culex portesi and Psorophora ferox (Map 20d). Epidemics occur regularly from Texas to Peru (Map 18d) affecting horses and people, the symptoms being severe and often fatal.4. TICK-BORNE DISEASES
M. G. R. Varma4.1 Tick-borne virus diseases
4.1.1 Colorado Tick fever (CTF)
Pathogen: Colorado Tick fever virus, Family Reoviridae, genus
Orbivirus
Vectors: Dermacentor andersoni, Family Ixodidae
Reservoirs: Various small mammals, particularly the golden mantled
ground squirrel (Spermophilus lateralis), Colombian ground squirrel
(Citellus colombianus), yellow pine chipmunk (Eutamias amoenus)
and least chipmunk (E. minimus).
A zoonosis acquired from a focal ecological niche. Foci exist in the Rocky Mountain states, South Dakota and in western Canada. In man it exhibits as a febrile illness with an exceptionally enduring viraemia and no skin rash, the latter differentiating it from Rocky Mountain spotted fever transmitted by the same species of tick. Although Dermacentor andersoni is the principal vector, the virus has been isolated from other species of ixodid ticks. Small mammals and immature D. andersoni maintain the infection in natural foci.4.1.2 Louping ill (Ll)
Pathogen: Louping ill virus, Family Togaviridae, genus Flavivirus
Vector: Ixodes ricinus, Family Ixodidae
Reservoirs: sheep, red deer (Cervus elaphus), red grouse
(Lagopus scoticus)
A biphasic meningoencephalomyelitis principally affecting sheep, but also cattle, red grouse, red deer and man in rough hill pasture in the British Isles. Ixodes ricinus is the only known vector; its typical biotopes are areas where the soil remains damp throughout summer, as in poorly drained hill pastures. The tick also occurs in forests in the South of England as well as on continental Eurasia, but is infected with louping ill virus only in Britain and Ireland. A variety of small mammals and ground frequenting birds serve as hosts of the immature tick. Sheep, red deer and cattle are the major hosts of adult ticks. Human cases occur among people with close contact with sheep and tick biotopes, e.g., shepherds and abattoir workers. Transmission to man is by bite of infected ticks. There is trans-stadial passage of virus but transovarial transmission has not been demonstrated.4.1.3 Encephalitides
Tick-borne encephalitis (TBE) (= Far Eastern or Russian Spring-Summer encephalitis (RSSE), Central European or Western tick-borne encephalitis (TBE), biphasic meningoencephalitis, diphasic milk fever)
Pathogen: TBE virus, Family Togaviridae, genus Flavivirus
Vectors: Ixodes ricinus (Western form, TBE); I. persulcatus
(Far Eastern form, RSSE), Family Ixodidae
Reservoirs: Small mammals; ticks through transovarial transmission
The Far Eastern form, often referred to as Russian Spring-Summer encephalitis (RSSE), is associated with the taiga forest and is transmitted chiefly by Ixodes persulcatus. This tick is intimately associated as larvae and nymphs with many small forest mammals and birds, and as adults with larger wild and domestic mammals. The tick requires 2-4 years to complete its life cycle. In western USSR and in eastern and western Europe, it is replaced by I. ricinus, the chief vector of the western type of tick-borne encephalitis. In the coniferous and temperate deciduous forests of eastern Europe extending from the Urals to the Atlantic Ocean, I. ricinus is the most common tick species. In western USSR, the ranges of the two species overlap and both types of the disease may occur. Other tick species may be involved to a minor extent and transmission is possible by drinking infected goats' milk as well as by tick bite.4.1.4 Kyasanur Forest disease (KFD)
Pathogen: KFD virus, Family Togaviridae, genus Flavivirus
Vectors: Ticks of the Family Ixodidae, principally Haemaphysalis
spinigera and H. turturis
Reservoirs: Possibly monkeys and small rodents
The disease was discovered in Kyasanur Forest in South India in 1957 following investigations of deaths of monkeys, Presbytis entellus and Macaca radiata in the forest and first suspected to be jungle yellow fever. Human infection with a high mortality in the initial outbreaks, was always associated with activities such as wood gathering and cattle grazing in the forests.4.2 Tick-borne borrelioses
4.2.1 Tick-borne relapsing fever (= tick-borne spirochaetosis, endemic relapsing fever)
Pathogen: Borrelia spp., about seven species cause disease in
man.
Vectors: Soft ticks of the Family Argasidae, genus Ornithodoros.
Reservoirs: Ornithodoros ticks, through transovarial transmission.
In the Old World, the disease is endemic in East, central and South Africa, the Mediterranean region (Portugal, Spain, North and West Africa) extending eastwards through Cyprus, Israel, Iran, Central Asia, Kashmir (India) to western China (Map 23, page 66). In the New World, it occurs in central and western USA, Middle and South America southwards to northern Argentina (Map 24, page 67). Several hundred human cases are reported annually. The mortality is seldom high, but morbidity may be severe. It is effectively treated with antibiotics.4.2.2 Erythema borreliosis migrans (= erythema chronicum migrans, Lyme disease)
Pathogen: Borrelia burgdorferi.
Vectors: Ixodes spp., I. ricinus in Europe, I. dammini in
north-eastern USA, I. pacificus in California and probably I. holocyclus
in Australia.
Amblyomma americanum is a possible vector in USA.
Reservoirs: Rodents and deer.
The disease was first recognized in Sweden in 1909 and later reported from Austria and other parts of continental Europe. It appeared in the USA in 1970 and caused an epidemic outbreak of arthritis in New England in 1972 in Old Lyme which led to the entity being named first Lyme arthritis then Lyme disease. In 1977 it was reported in the United Kingdom and in 1982 in Australia (Map 25, page 68).4.3 Tick-borne rickettsioses
4.3.1 Rocky Mountain spotted fever (= RMSF, Mexican spotted fever, Tobia fever, Sao Paulo fever)
Pathogen: Rickettsia rickettsi.
Rocky Mountain spotted fever is a zoonotic infection widely distributed in the Nearctic and Neotropical regions from Canada to South America, now mostly prevalent in eastern USA where the mortality and morbidity rates remain unacceptably high (Map 26, page 69). With the advent of antibiotics, the mortality rate in human beings has been reduced. In parts of eastern USA, with an increasing human population and the encroachment of residential areas into abandoned fields and woodland with considerable rodent and tick activity, the attack rate is high and women and children account for at least half of the reported cases; most cases in eastern and southern USA are rural and suburban. Apart from the three species of ticks mentioned above, others, particularly Rhipicephalus sanguineus have been found naturally infected, and may be involved in the maintenance of natural foci of infection and the transmission of infection to man.4.3.2 Boutonneuse fever (= South African tick typhus, Kenya tick typhus, Marseilles fever, Crimean tick typhus, Indian tick typhus); Siberian tick typhus (= North Asian tick typhus); Queensland tick typhus
Pathogen: Rickettsia conori(Boutonneuse fever), R. sibirica
(Siberian tick typhus), R. australis (Queensland tick typhus).
Vectors: Ticks of the Family Ixodidae.
Reservoirs: Ticks and rodents.
Boutonneuse fever is known by various local names and is widely distributed in Africa, including Egypt, where it was recently found. It also occurs in the European and African areas near the Mediterranean and in Israel, and over much of southeastern Asia. The name is derived from the button-like lesion at the site of attachment of the tick.5. MITE-BORNE RICKETTSIOSIS
M.G.R. VarmaChigger-borne rickettsiosis (= Tsutsugamushi disease, Japanese river fever, scrub typhus, mite-borne typhus)
Pathogen: Rickettsia tsutsugamushi (= R. orientalis).
Vectors: Mites of the Family Trombiculidae, Genus
Leptotrombidium
Reservoirs: Mites, through transovarial transmission; various spp.
of rodents, especially Rattus.
A zoonotic infection, first described from Japan, now known to occur over most of southeastern Asia, India, Sri Lanka, Pakistan, islands of the southwest Pacific, and coastal Queensland (Australia) (Map 28). It is found in a wide range of biotopes from flooded alluvial plains in Japan, scrub, and dense but disturbed forest in southeastern Asia, to semi-deserts as in Pakistan and alpine reaches in the mountains as in the Himalayas. The actual distribution may be much wider. A high incidence occurs in human beings exposed to infected biotopes. There were large outbreaks in nonimmune troops in southeastern Asia during the Second World War and the Vietnam War, with a high mortality.6. LOUSE-BORNE DISEASES
J. Maunder
Pathogens: Rickettsia prowazeki, (classical typhus)
Rochalimaea quintana (trench fever, now sometimes called
quintana fever)
Borrelia recurrentis (louse-borne relapsing fever, a spirochaetal
disease, to be distinguished from the clinically similar but
epidemiologically distinct tick-borne relapsing fever caused by
Borrelia duttoni, from which it may be derived.)
Vectors: Pediculus humanus humanus (the human body louse,
exclusively incriminnated as a vector of endemic typhus and trench
fever). Pediculus humanus capitis (the head louse) has no practical
importance as a vector. Pthirus pubis (the crab louse) is neither a
disease vector but infestations of this species are often associated with infections with sexually transmitted diseases.
Lice are also thought by some to be mechanical vectors of bacterial
skin infections, especially of impetigo and staphylococcal boils.
Persistently louse-infected individuals may develop a malaise
directly attributable to the lice and allergy to louse faeces has also been
reported.
All three human lice are cosmopolitan, but disease transmission is linked with the prevalence of the body louse (Pediculus humanus humanus), the principal vector. This louse is favoured when people live under conditions of cold, crowding, poverty or lack of hygienic facilities, especially when there is competition for insufficiently available clothing .7. TRYPANOSOMIASIS
7.1 African (Sleeping Sickness)
D. M. Minter
Pathogens: Trypanosoma (Trypanozoon) brucei gambiense in West
Africa; T. (T.) brucei rhodesiense in East, central and southern
Africa.
Vectors: Tsetse flies (Diptera: Glossinidae: genus Glossina).
Reservoirs: Recent work suggests that T. (T.) b. gambiense has a
mammalian reservoir other than man, which may be important for
the persistance of foci, but further studies are needed (WHO, 1986).
T. (T.) b. rhodesiense is fundamentally a parasite infecting
ruminants which are the principal reservoir hosts.
Sleeping sickness occurs at a low level of transmission in the majority of countries in tropical Africa, with occasional epidemic outbreaks and at least 50 million people are at risk (Map 31a). Animal trypanosomiasis deters the breeding of domestic stock over large areas of the continent (WHO, 1979).7.2 American (= Chagas disease)
C. J. Schofield
Pathogen: Trypanosoma (Schizotrypanum) cruzi
Vectors: Triatomine bugs (Triatoma). Houseflies and cockroaches
may act as carriers
Reservoirs: Rodents, marsupials and man
Chagas disease and its vectors are distributed throughout the American continent and some Caribbean islands, roughly between latitudes 40\'a1N and 46\'a1S (Map 33). Some potential triatomine vectors also occur in Africa, Asia and Australia, but the causative agent, Trypanosoma cruzi, has not been reported from the Old World (Map 33).8. THE LEISHMANIASES
D. M. Minter
Pathogens: The genus Leishmania contains two subgenera: the
subgenus Leishmania Saf'janova 1982 and the subgenus Viannia
Lainson & Shaw 1987. 1
Leishmania (Leishmania) donovani and L. (L.) infantum are the
causative organisms of visceral leishmaniasis (VL), or kala-azar,
in the Old World and L. (L.) chagasi is the New World VL parasite.
Leishmania (L.) aethiopica, L. (L.) major and L. (L.) tropica are
the causative organisms of cutaneous leishmaniasis (CL) in the Old
World. The parasites which cause cutaneous leishmaniasis in the
New World are more numerous and include the L. (L.) mexicana
complex of species [L.(L.) amazonensis, L. (L.) mexicana and L. (L.)
venezuelensis] and two species possibly also of the L. (L.) mexicana
complex [L. (L.) gamhami and L. (L.) pifanoi].
Other CL parasites of the New World are species of the subgenus
Viannia: most belong to the L. (V). braziliensis complex [L. (V.)
braziliensis, L.(V.) quyanensis, L. (V.) panamensis and L. (V.)
peruviana]. Leishmania which commonly cause human diffuse
cutaneous leishmaniasis (DCL) are L.(L.) aethiopica (in the Old
World); L. (L.) amazonensis, L. (L.) pifanoi and an unnamed
Leishmania (L.) sp. (in the Dominican Republic) are particularly
associated with human DCL in the New World.
The Leishmania species most closely associated with human
mucocutaneous leishmaniasis (MCL) is L.(V.) braziliensis (in the New
World: the cause of espundia). Other Leishmania more occasionally
cause mucocutaneous lesions; e.g. in the Sudan, where an ill-defined
Leishmania causes MCL (the organism is sometimes given specific
status as L.(L.) archibaldi, but it is very close to, or possibly
identical with, L.(L.) donovani).
Vectors: Phlebotomus spp. in the Old World and Lutzomyia spp.
in the New World.
Reservoirs: VL - man, dogs, wild carnivores
CL - man (L.(L.) tropica); rodents (L.(L.) major); hyrax (L.(L.)
aethiopica); sloths, opossums, etc. (L.(V.) braziliensis complex);
rodents (L.(L.) mexicana complex).
The parasites that cause leishmaniasis are a morphologically similar, but biologically diverse, group of organisms which are responsible for a wide spectrum of human disease. Most leishmanial diseases are zoonoses, with a wide variety of natural animal hosts that normally are little affected by the presence of the parasites.
(Ph.) = Phlebotomus
(Pa.) = Paraphlebotomus
Sy.) = Synphlebotomus
(La.) = Larroussius
(Ad.) = Adlerius
(Eu.) = Euphlebotomus
(Lu.) = Lutzomyia
(Pt.) = Pintomyia
(Ny.) = Nyssomyia
(Ps.) = Psychodopygus
REFERENCES9. FLEA-BORNE DISEASES
I.A.H. Ismail and J. Maunder9.1 Plague
Pathogen: Yersinia (Pasteurella) pestis
Vectors: Fleas
Reservoirs: Rodents
Plague is a zoonotic disease, primarily of rodents. Human involvement is basically opportunistic. The disease is transmitted from rodent to rodent and from rodent to man by fleas. Man can also acquire the infection through direct contact with infected animal tissues. Pneumonic plague can result from direct human to human transmission.9.2 Murine typhus
Pathogen: Rickettsia mooseri
Vectors: Fleas
Reservoirs: Rattus spp.
Murine typhus, also called flea-borne typhus is of cosmopolitan distribution, this rickettsiosis occurs in mini-foci in buildings, especially in port cities in the subtropics and tropics (Map 47).10. ONCHOCERCIASIS
Pathogen: Onchocerca volvulus - river blindness, onchocerciasis,
Robles' disease (Guatemala), sowda (Yemen)
Vectors: Blackflies of the genus Simulium (Order Diptera,
Family Simuliidae)
Reservoirs: Endemic mainly in man, O. volvulus has also been
found naturally in the gorilla (Gorilla) in Zaire and it can be
transmitted experimentally to the chimpanzee (Pan), but the zoonotic
reservoir is unimportant in relation to human disease foci which always
seem to be sustained exclusively by man-to-man transmission.
Onchocerciasis occurs focally in tropical America, from southern Mexico southwards to the equator (Map 48), and more extensively in tropical Africa, with a small focus in Arabia overlapping the borders of Yemen and Saudi Arabia (Map 49). Human infection rates and symptoms vary greatly among the foci owing to variations in the form or strain of O.volvulus as well as the local characteristics of vectors and human exposure patterns (WHO, 1987). The most pathogenic form of onchocerciasis is widespread across West African savannas, where a high proportion of adults in some communities have been blinded by chronic infection.11. LOIASIS
Pathogen: Loa loa
Vector: Chrysops spp.
Reservoir: Man
Loiasis, a filarial disease affecting about one million people, is limited in distribution to forest areas in Central Africa from eastern Nigeria to Angola, extending into Congo, Zaire, Uganda and the Sudan (Map 54) (Rhodain, 1980).
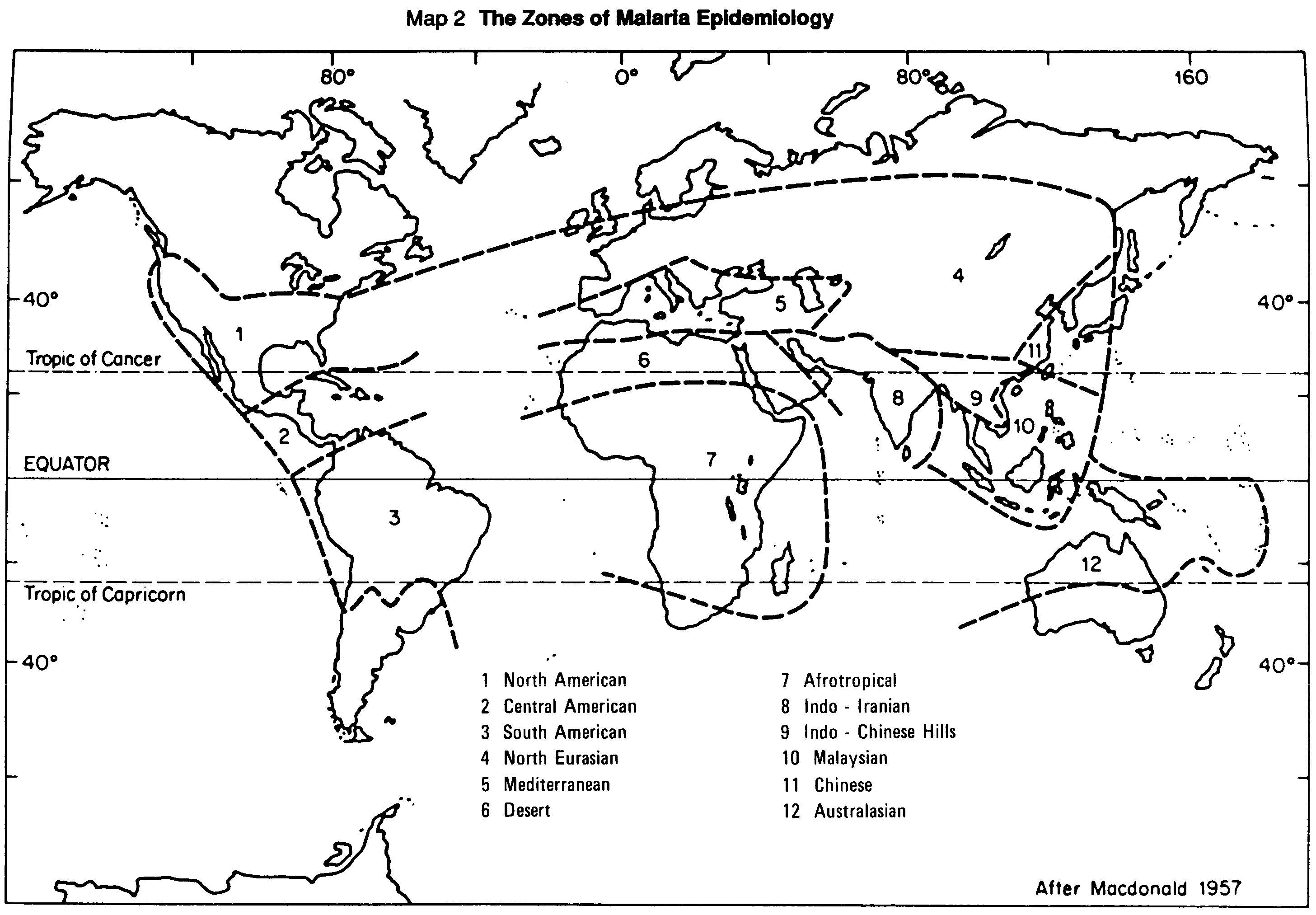{kind=link}
{kind=link}
{kind=link}
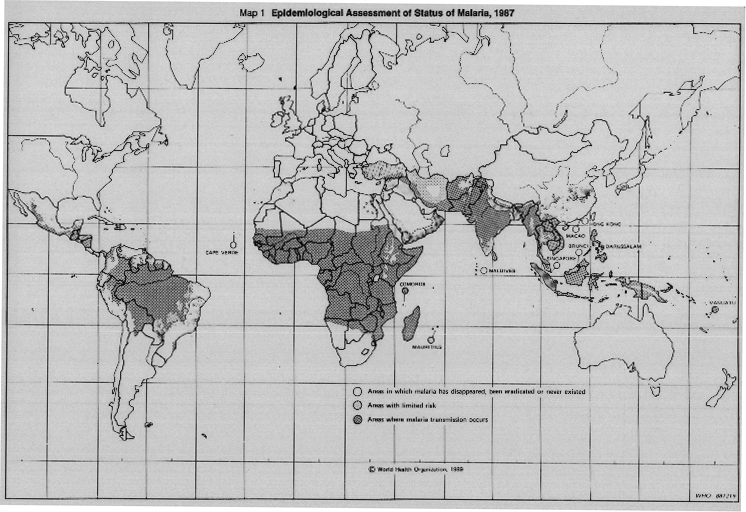{kind=link}
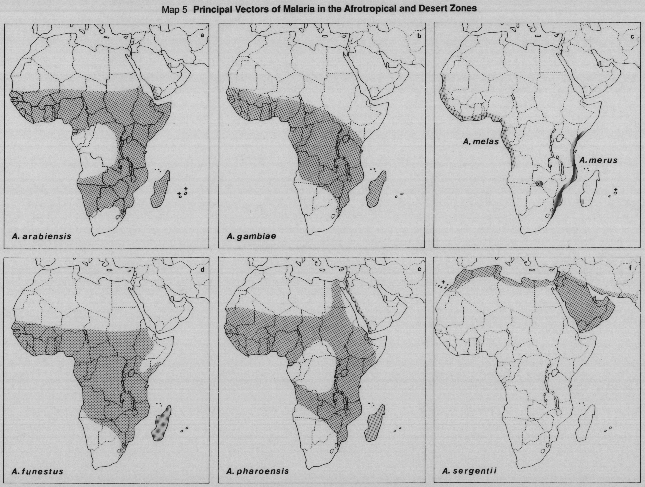{kind=link}
{kind=link}
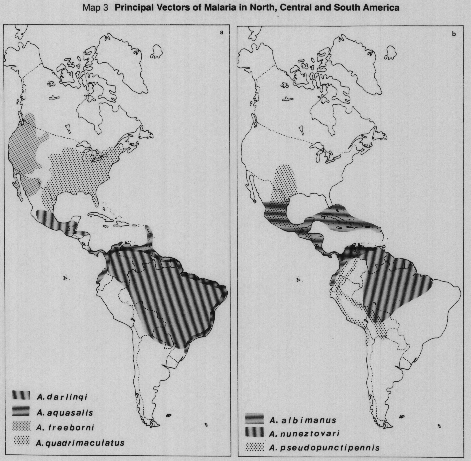{kind=link}
{kind=link}
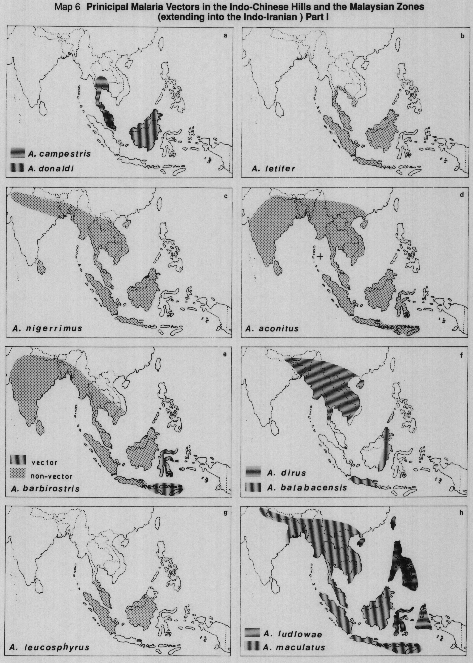{kind=link}
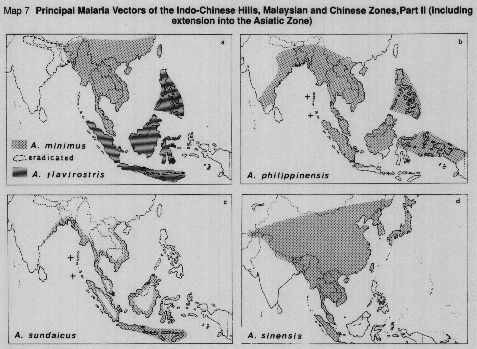{kind=link}
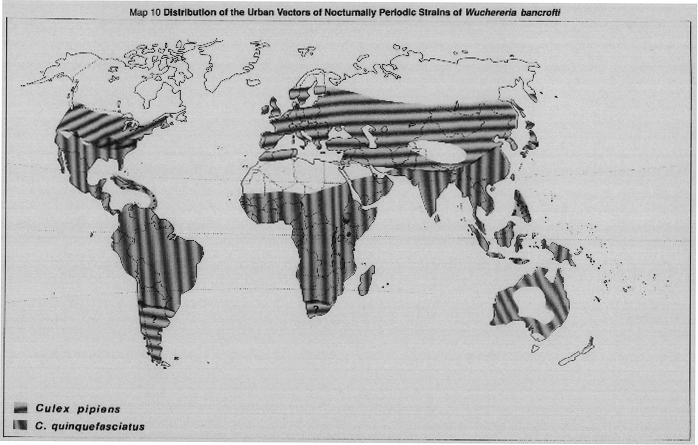{kind=link}
{kind=link}
{kind=link}
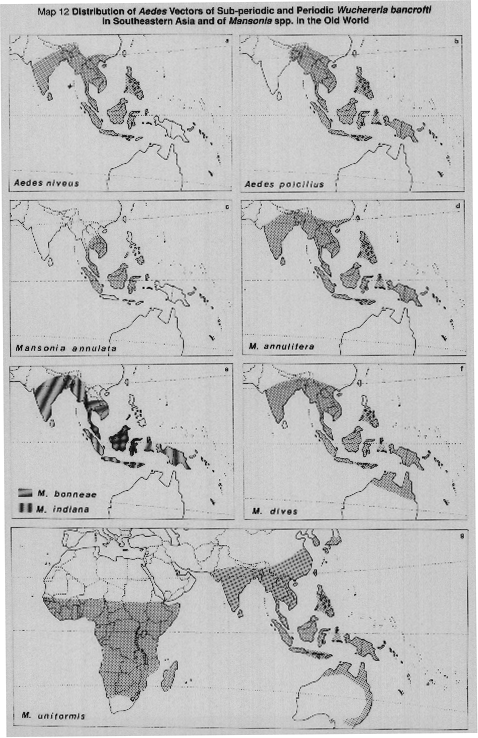{kind=link}
![[a]](tab2a.gif){kind=link}
![[b]](tab2b.gif){kind=link}
![[c]](tab2c.gif){kind=link}
![[d]](tab2d.gif){kind=link}
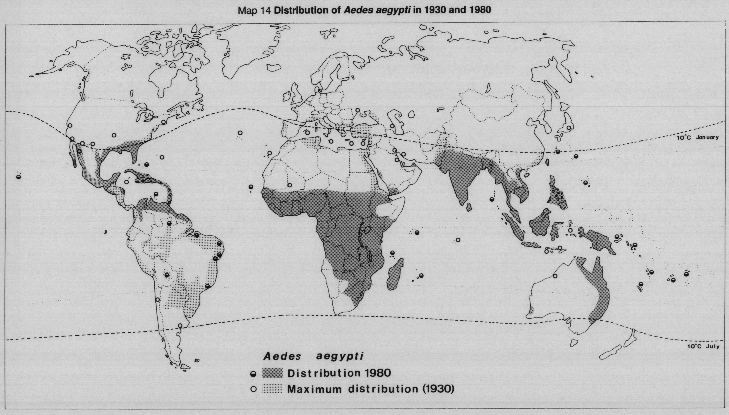{kind=link}
{kind=link}
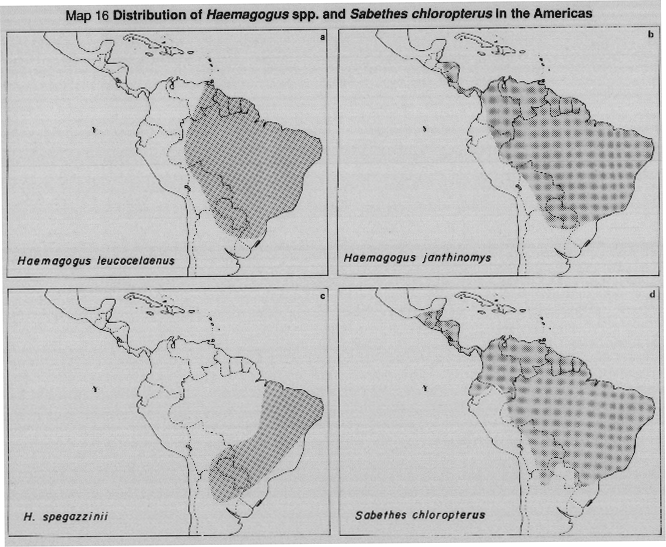{kind=link}
{kind=link}
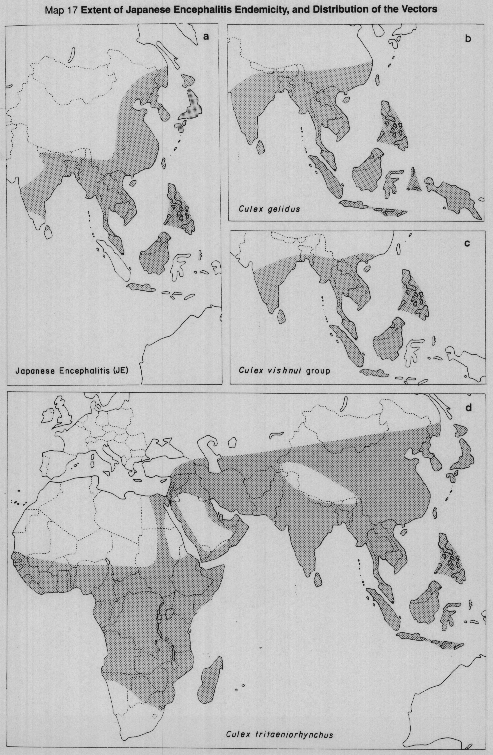{kind=link}
{kind=link}
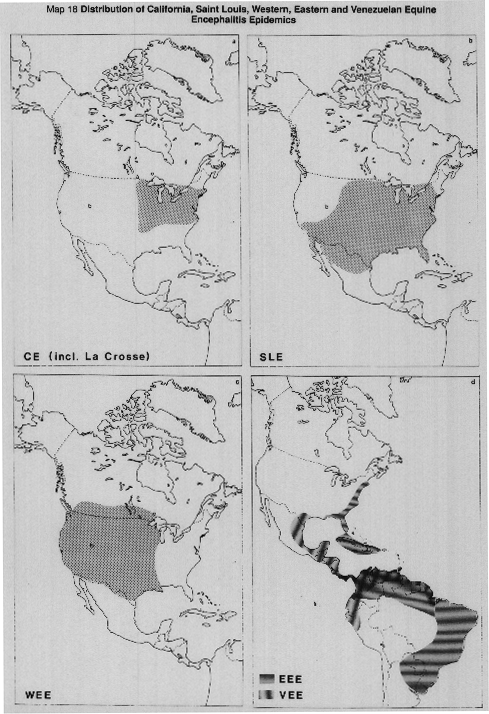{kind=link}
{kind=link}
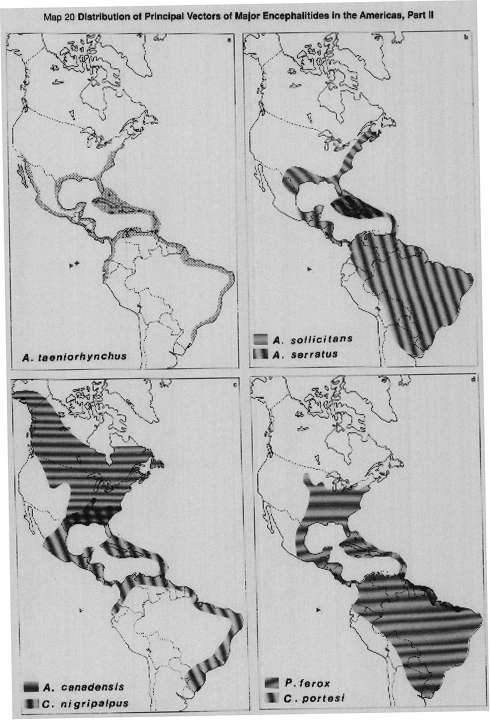{kind=link}
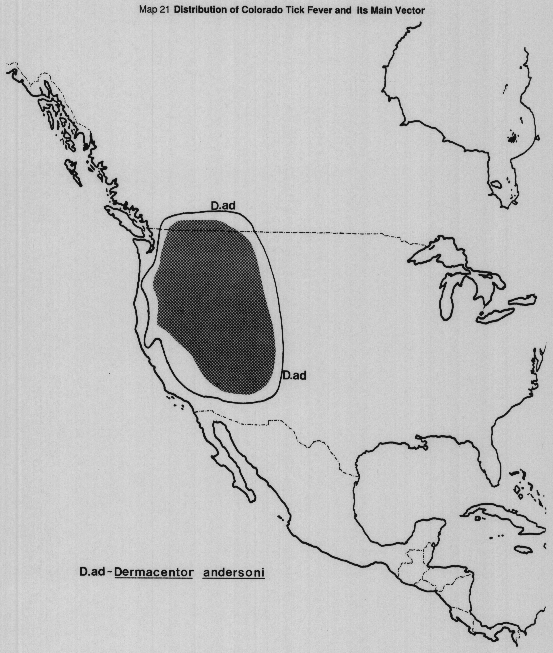{kind=link}
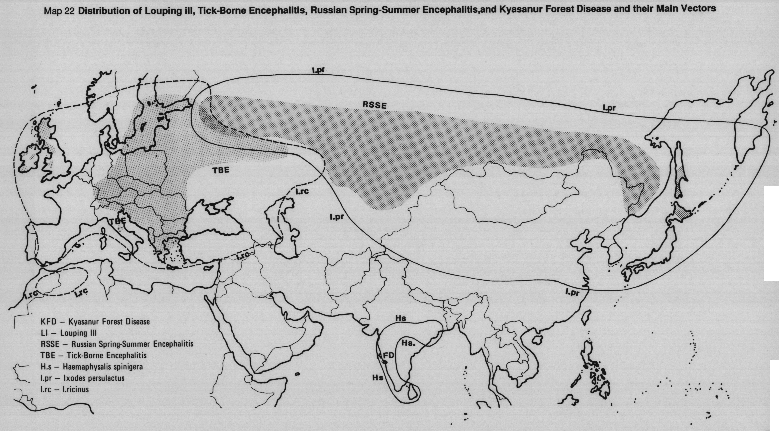{kind=link}
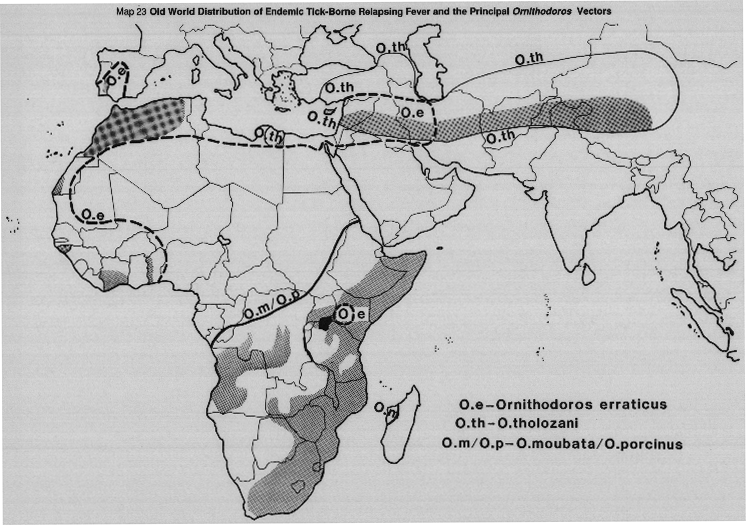{kind=link}
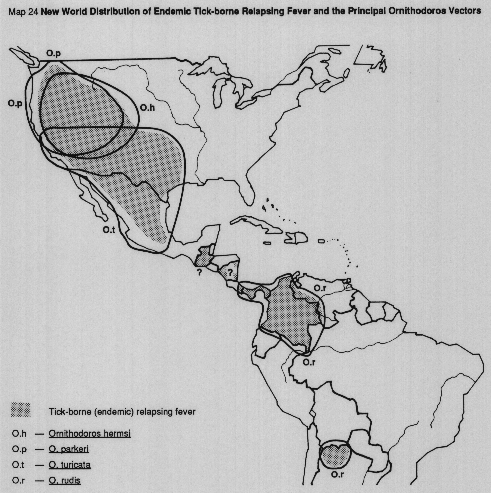{kind=link}
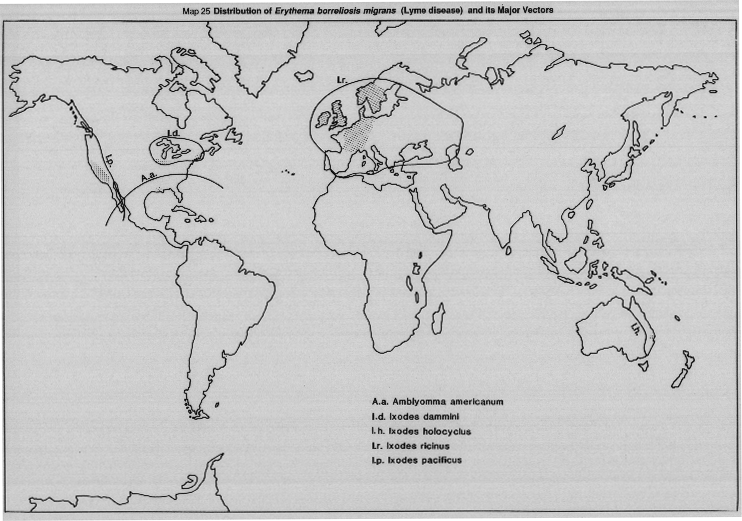{kind=link}
{kind=link}
{kind=link}
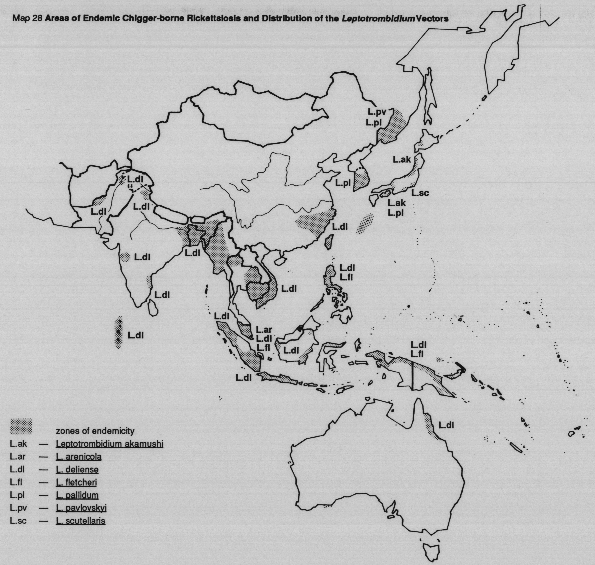{kind=link}
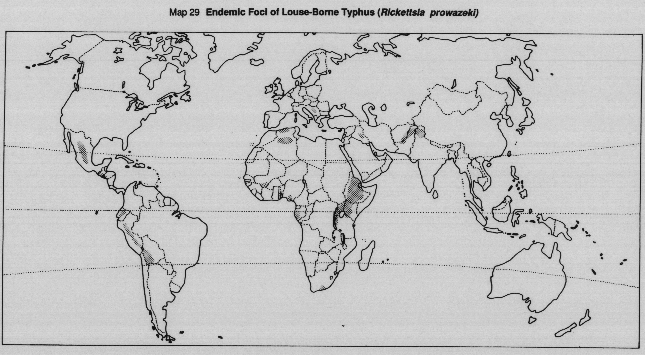{kind=link}
{kind=link}
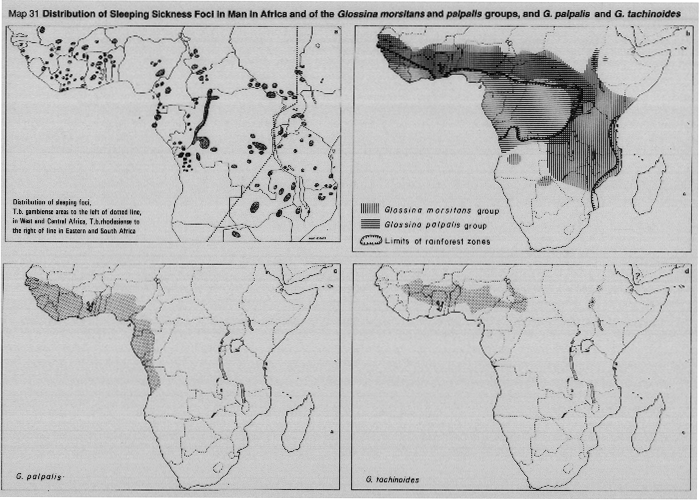{kind=link}
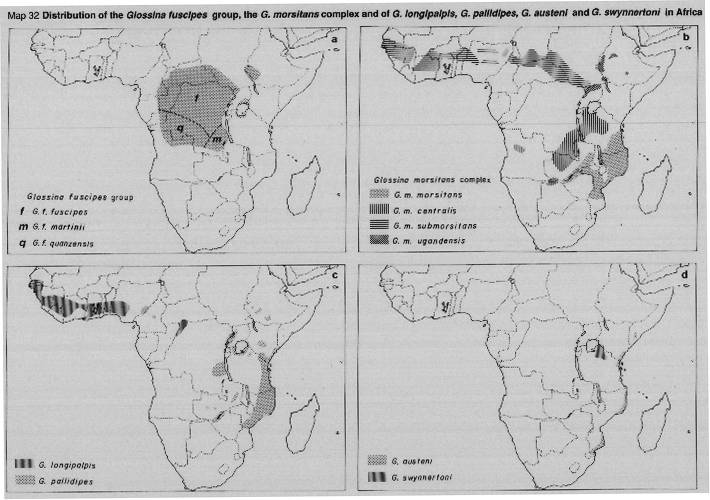{kind=link}
{kind=link}
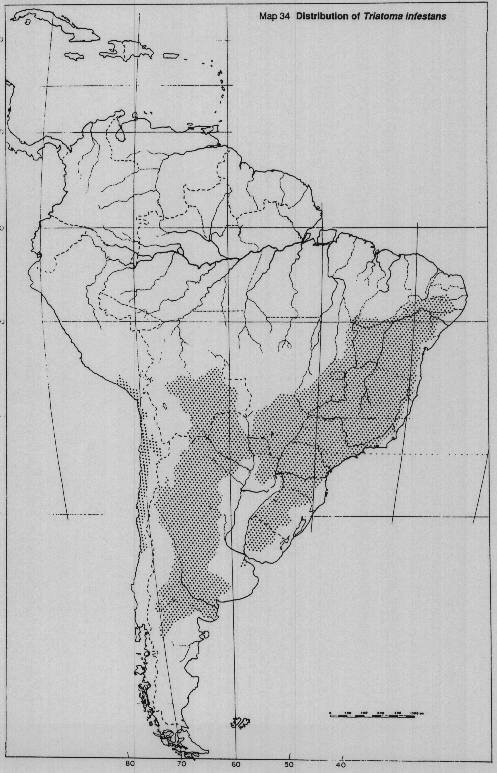{kind=link}
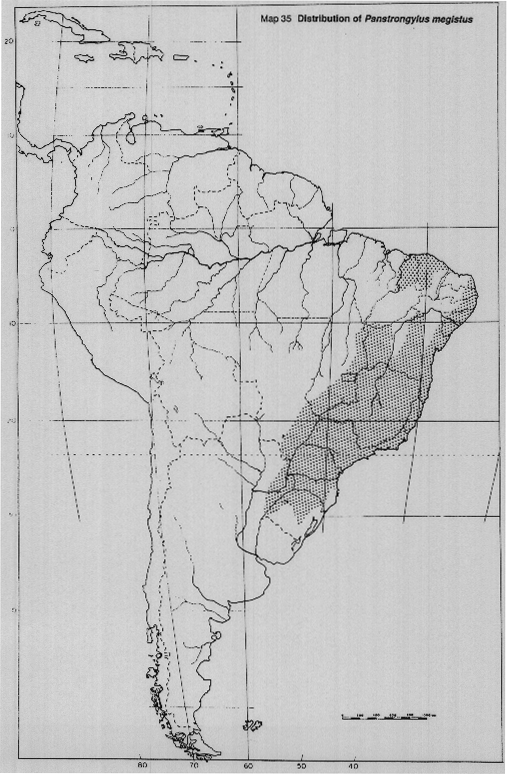{kind=link}
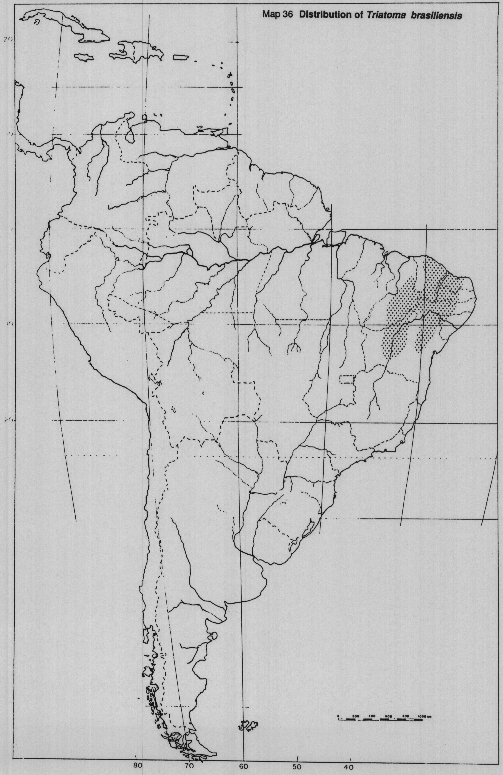{kind=link}
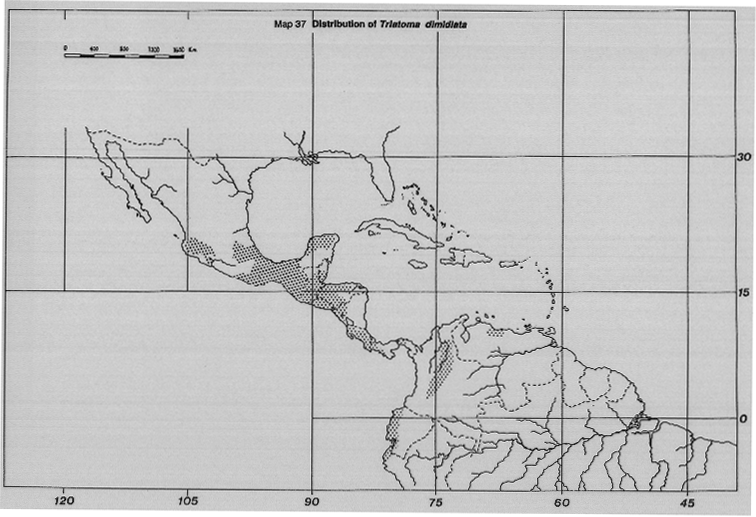{kind=link}
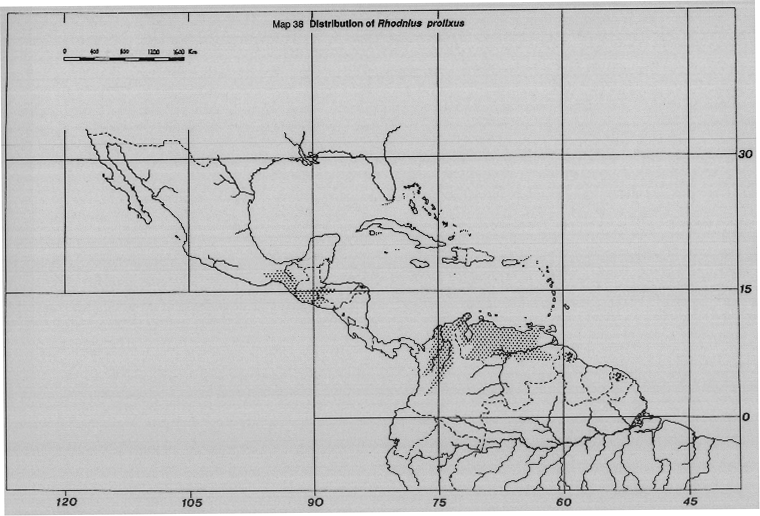{kind=link}
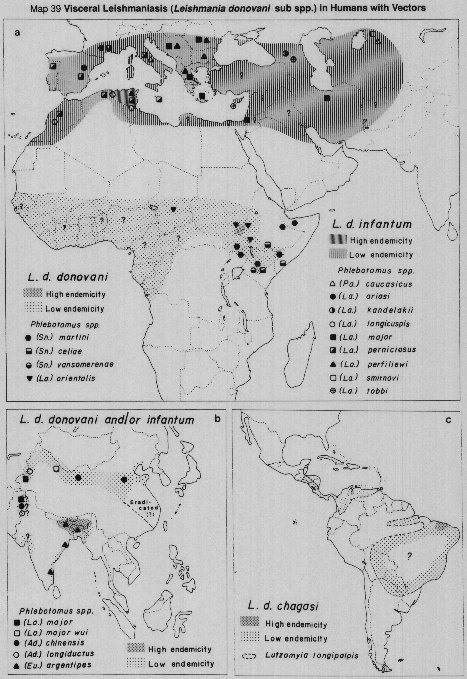{kind=link}
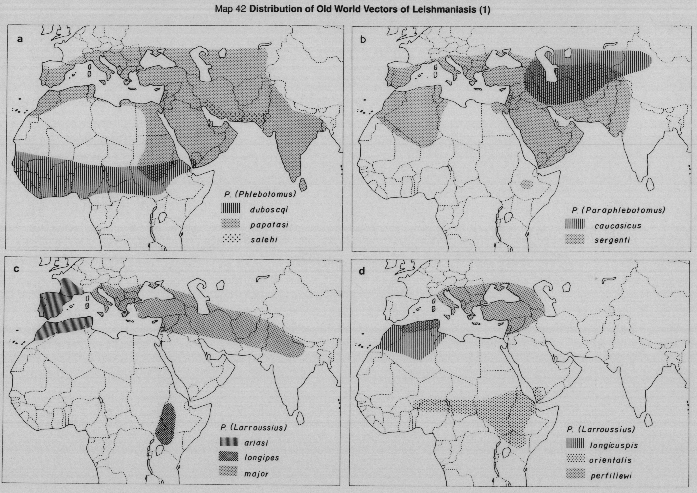{kind=link}
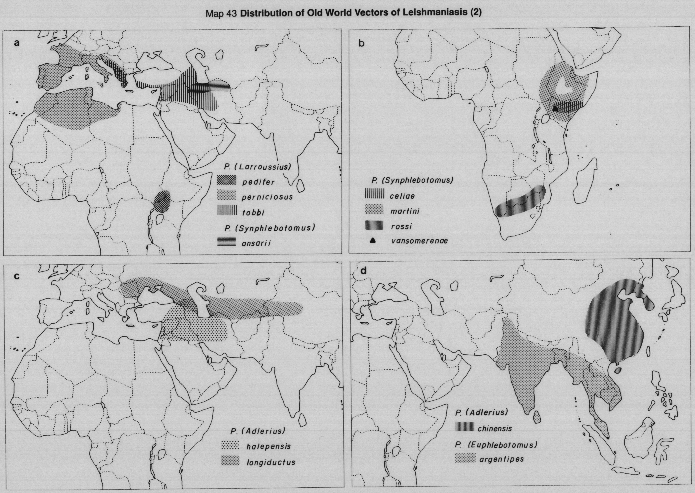{kind=link}
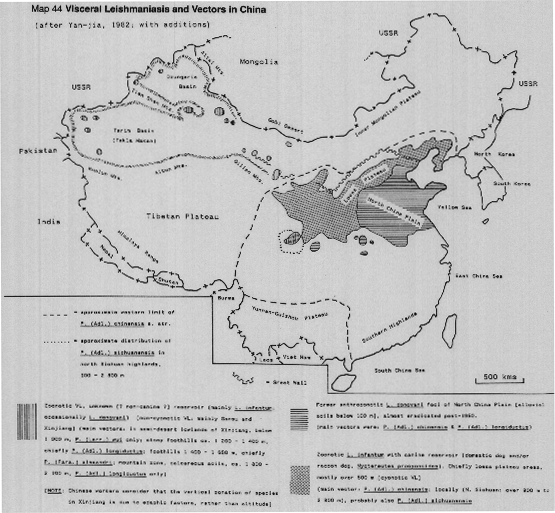{kind=link}
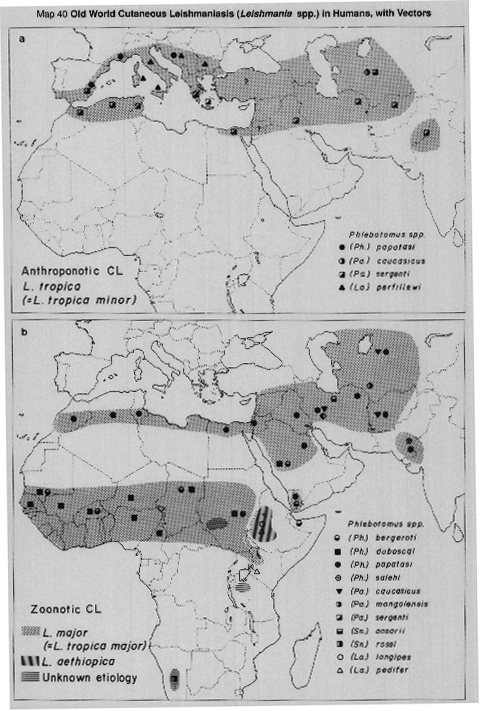{kind=link}
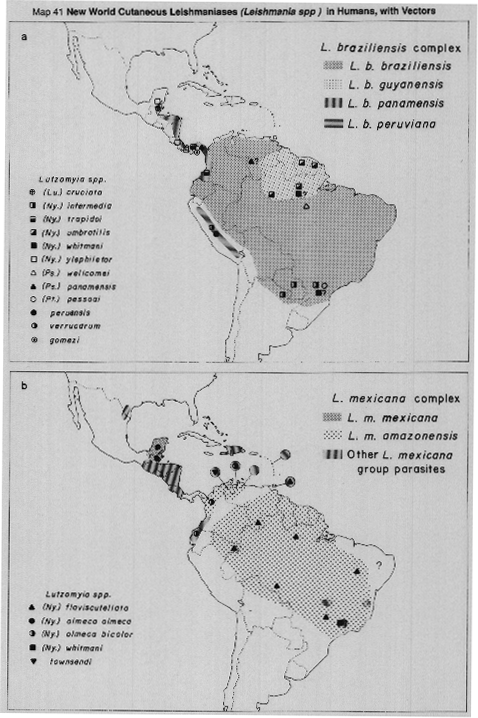{kind=link}
{kind=link}
![[Appendix]](tab100.gif){kind=link}
{kind=link}
{kind=link}
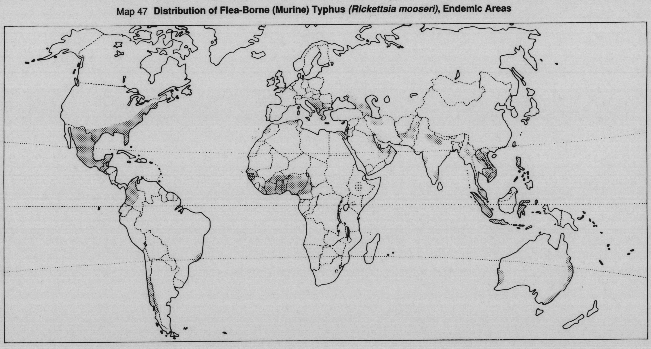{kind=link}
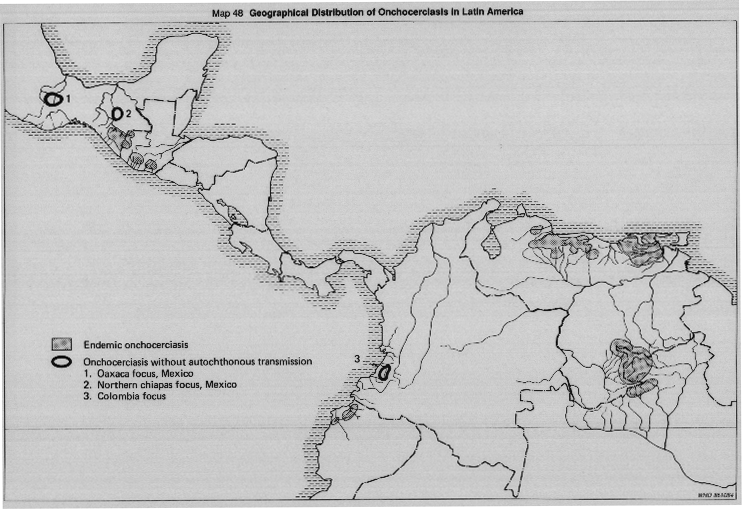{kind=link}
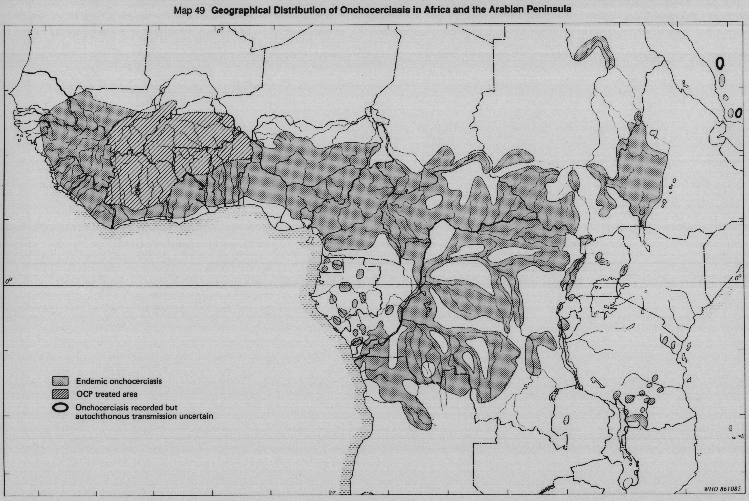{kind=link}
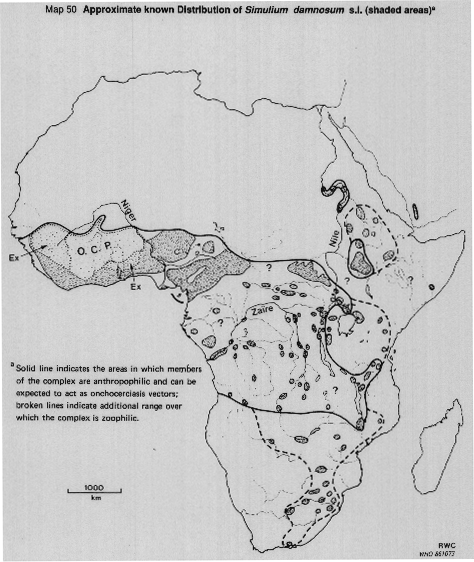{kind=link}
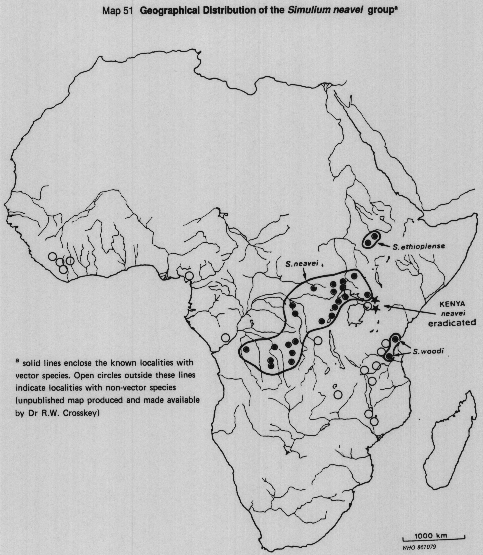{kind=link}
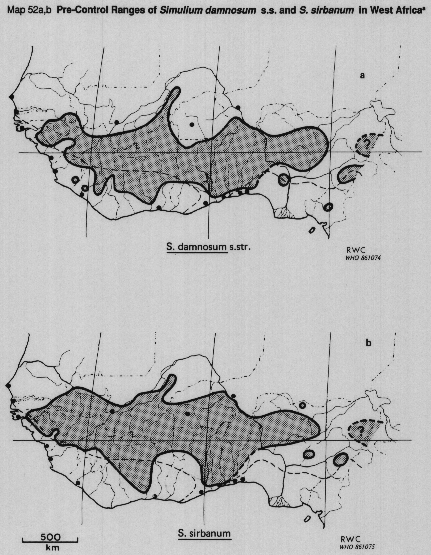{kind=link}
{kind=link}
{kind=link}
{kind=link}
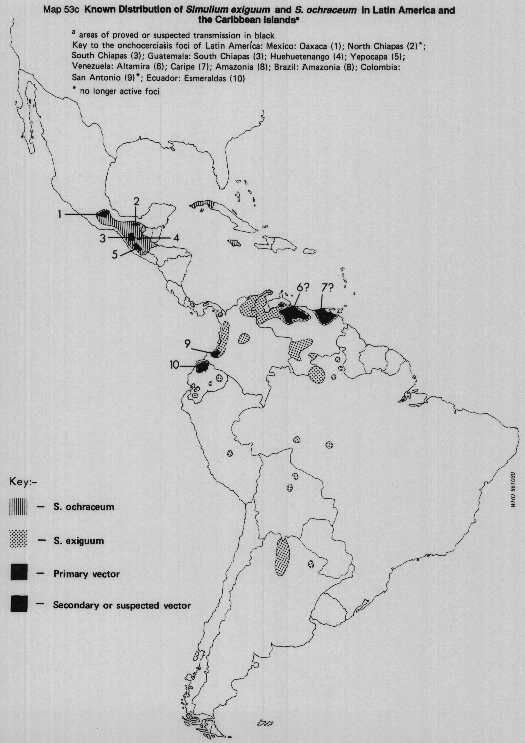{kind=link}
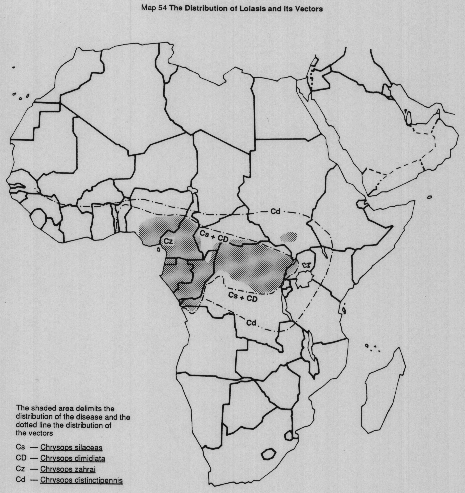{kind=link}