 Reproduced, with permission, from:
Leng, R. A. 1993. Quantitative ruminant nutrition - A green science. Australian Journal of Agricultural Research 44: 363-80.
Reproduced, with permission, from:
Leng, R. A. 1993. Quantitative ruminant nutrition - A green science. Australian Journal of Agricultural Research 44: 363-80.
Department of Biochemistry, Microbiology and Nutrition, University of New England, Armidale, N.S.W. 2351.
Abstract
Knowledge of quantitative digestion and metabolism in ruminants was developed most rapidly when isotope dilution techniques became easy to apply, facilitated by improved instrumentation and mathematical approaches. The Armidale group led by Professor E. F. Annison and Dr. D. B. Lindsay were at the forefront of these developments in the late 1950's. Since then knowledge in this area has developed at an ever increasing rate. The data that accumulated from the quantitative approach led to simple or complex models of animal digestion, metabolism and growth. These in turn led to much questioning of the dogma of feed evaluation and feeding standards as they applied in practice, especially for ruminants fed on poor quality forages. The knowledge that developed has clearly shown that the way toward substantial increases in productivity of ruminants on forage based diets is through the balanced nutrient approach that considers the efficiency of the rumen ecosystem and the availability of dietary nutrients post-ruminally. With increasing emphasis on quality-beef markets at the present time, it seems likely that the time is ripe for application of much of this knowledge. The major breakthroughs have come about by recognition of the nutrients required to balance a ruminant's diet where the animal depends on the end-products of rumen fermentation (i.e. on a forage-based diet). When this is achieved, the increase in efficiency of use of nutrients lifts the overall nutrition of the animal to a level that is well above that predicted from feeding standards, based on the metabolizable energy content of the supplement or the total diet. This understanding, together with the stoichiometry of rumen fermentation, has indicated an important approach to help ameliorate the greenhouse effect, that is, lowering of enteric methane production per unit of feed intake or per unit of animal products from ruminants by strategic supplementation.
Keywords: methane production, greenhouse effect, feed conversion efficiency.
The early research centered initially on analytical technology and the major questions related mainly to the digestive mode of the ruminant: what nutrients were important in their metabolism and how did they rank in quantitative importance (Annison et al. 1967). At that time debate raged over whether glucose was important in ruminant metabolism (Annison and White 1961) and whether ketone bodies were normal substrates or toxic by-products of fat metabolism. The period was exciting. As an example of the lack of knowledge of the quantitative contribution of circulatory nutrients to the nutrition of ruminants in the early 1960's, the following example is given: when radioactivity labelled B-hydroxybutyrate was first prepared by incubating (l4C)-butyrate with sheep liver slices in vitro (Leng and Annison 1963), the opportunity arose for the first time to examine ketone body metabolism in the whole animal (Leng and Annison 1964). In the first run in which labelled B-hydroxybutyrate was infused into a mature sheep and both B-hydroxybutyrate and carbon dioxide isolated for assay of specific radioactivity, it was estimated that over 80% of the carbon dioxide produced by the sheep arose from oxidation of B-hydroxybutyrate (Leng, R. A., unpublished observations). This was hailed as a major breakthrough in knowledge; however, it was soon realized that a rather simple analytical error had created an enormous error in the estimate. The specific radioactivity of the extracted B-hydroxybutyric acid had been estimated in a two step process. The radioactivity of B-hydroxybutyrate was estimated after evaporating the extracting solvent without prior conversion to a salt; a proportion of the B-hydroxybutyrate apparently volatilized with the solvent. The concentration of the acid had, however, been determined by titration of a separate portion. The result was a very low apparent specific radioactivity of the B-hydroxybutyrate in blood and an extremely high estimate of its entry rate. Repeating the experiment with neutralization of the solvent prior to evaporation put B-hydroxybutyrate into perspective as a minor nutrient in ruminant metabolism.
The early technology development and surveys of the contribution of nutrients to oxidative metabolism (Annison et al. 1967) gave way to the use of these technologies to quantify differences between animals on different diets. This led ultimately to their use in research aimed at problem solving on a local and global scale.
Those of us who have attempted to apply knowledge of quantitative digestion and metabolism to the establishment of feeding systems in the real world soon recognized that these assumptions do not, generally, apply; in particular, they do not apply to the 'forage fed' ruminant. Under production systems, nutrient availability to the rumen microbes often sets the upper limit of production through a low microbial growth efficiency in the rumen (which directly results in a low protein to energy ratio in the nutrients absorbed). The other limitation is the amount of dietary protein that escapes rumen digestion, but which is digested in the intestines. Ruminants require more protein generally (particularly in tropical climates) than arises from microbial growth in the rumen whether the latter is efficient or inefficient (Leng 1990a).
The majority of the world's ruminants depend over their lifetime on forages that can be described only as poor quality, in which the limitations to production are a low protein supply from the microbial ecosystem and a virtual absence of dietary bypass protein.
The efficiency of microbial growth, in the rumen (expressed as g cells produced per ATP, or per mole of VFA, or per g of carbohydrate fermented) appears to be highly variable depending on the feeding conditions. This variability must be much greater in the field than is generally observed in the controlled laboratory studies. For example, responses under field conditions to supplements aimed largely at correcting nutrient deficiencies of the rumen using multi-nutrient blocks (see Kunju 1988; Habib et al. 1991; Hendratno et al. 1991, and Saadulah 1991) are often large and are widespread through diverse feeding systems (see Loosli and McDonald 1968).
The factors that affect microbial growth efficiency and therefore the protein relative to VFA available for digestion and absorption are:
1. A deficiency of any microbial growth factor (e.g. ammonia, sulfur, phosphorus, amino acids, etc.) in the feed or induced some time after feeding in rumen liquor because of rapid absorption of the nutrient. For example, ammonia levels decrease rapidly after feeding to reach deficiency levels after 12 h on straw based diets, even if these are supplemented with urea.
2. The relative amounts of carbohydrate and protein that are fermented (fermentative degradation of protein and amino acids is coupled to a lower (approx. half) ATP generation than equivalent quantity of carbohydrate). A high protein to carbohydrate ratio in the diet can lead to a relatively low microbial protein to VFA ratio in the end-products of fermentative digestion where the dietary protein is easily and rapidly fermented in the rumen.
From the literature, the evidence seems to suggest that, if there are no deficiencies of microbial growth factors in the rumen, the source of the carbohydrate (sugar v. starch v. fibre) and the flow rate of liquor from the rumen appear to have little influence on microbial growth yield (see, for example, Maang Chang et al. 1989).
Bacterial lysis may occur from death of bacteria when their substrate in the rumen is exhausted (Hespell 1979). Lysis under these conditions may be triggered by temperate lytic phages that already infect rumen bacteria (see Klieve et al. 1989). Degradation in situ of rumen bacteria also results through predation of bacteria by protozoa (Coleman 1975). Lysis of bacteria in the rumen can severely reduce protein to energy ratio in the nutrients delivered to the animal for digestion and absorption (Leng and Nolan 1984). It is emphasized here that although, at times, microbial growth efficiency in the rumen can be high, the ratio of protein relative to energy that is available for digestion and absorption can be reduced markedly and can be extremely low due to this lysis. Nolan and Leng (1972), Mathieson and Milligan (1972), and Nolan and Stachiw (1979) demonstrated using stable isotopes of nitrogen that 25-50% of the microbial protein pool in the rumen may be turning over in situ and therefore unavailable.
Although there have been numerous attempts to measure microbial and total protein entering the duodenum of ruminants on various dietary regimes, the large errors associated with such measurements have precluded a systematic examination of the factors that affect both microbial growth efficiency and the protein entering the duodenum relative to the VFA produced in the rumen and absorbed. Research with animals cannulated at the rumen and duodenum and using markers have rarely given an accurate picture of the relationship between the end-products of fermentation (microbial cells entering the duodenum and VFA production). In some studies microbial growth in the rumen measured by these techniques have been at twice the theoretically possible rate, indicating that these technologies are very inaccurate and sometimes biased. In reviewing the literature, however, Czerkawski (1986) found values for microbial growth under a wide variety of feeding conditions to be very constant between studies.
Estimates of Y-ATP (g of dry microbial cells produced per unit of ATP theoretically generated in VFA formation) suggest that under good rumen conditions Y-ATP may be of the order of 10-14. Data collected from in vitro studies suggest that Y-ATP can be extremely low on diets where key microbial nutrients are deficient (perhaps as low as Y-ATP 4-5) and that with good conditions and when the protozoa are eliminated from the rumen ecosystem this could go as high as Y-ATP 20 (see Veira et al. 1983). These growth efficiencies would result in protein to energy ratios in the nutrients presented for absorption of, less than 5 to 20-30 g protein MJ of VFA energy respectively, indicating the scope for improvement in P:E ratios is high and therefore feed utilization improvements may be enormous (see later).
Although considerable responses are recorded to supplements of digestible protein that escapes degradation in the rumen in ruminants on all diets, the responses to animals on fibrous feeds have been the most dramatic. This is discussed in more detail below.
Understanding of the stoichiometry of the rumen and the requirements for additional feed protein that provide amino acids directly to the animal has allowed a new approach of establishing feeding strategies using locally available feeds. Essentially this is as follows:
This approach is now being applied on a large scale in many parts of the world. It suffers criticism because it does not depend on feed analysis which then becomes largely irrelevant (except for N estimation) in establishing feeding systems. It relies on 'rules of thumb' that require a depth of appreciation of the science of ruminant nutrition for their effective application A more scientific approach must eventually emerge, but at the present time we have no way of predicting the microbial growth efficiencies or the balance of nutrients absorbed by ruminant animals on any diet.
A major contribution to knowledge and production would be made by research that allowed ease of prediction of:
Armed with such a tool a considerable number of low productivity syndromes in practice could soon be understood.
When the two 'rules of thumb' concerning supplementation to achieve optimal microbial growth and for supply of protein post ruminally are applied to diets based on relatively poor quality roughages, surprisingly high levels of production can be achieved. For a diversity of research results that support this concept, see Hennessy et al. 1989; Perdok and Leng 1989; Silva et al. 1989; Hennessy and Williamson 1990 and Leng 1990b.
Ruminants contribute approximately 18-20% of the global methane produced annually (Gibbs et al. 1989). Enteric methane emission is one of the few global sources of methane that can be relatively simply reduced. It is more easy to manipulate than, for instance, methane produced from marshes or in rice production. Methane accumulation in the atmosphere requires only a slowing of emissions by 15-20% for world atmospheric concentration to stabilize.
A brief overview of the greenhouse effect is probably needed at this point in the discussion in order to present the link between quantitative nutrition and the strategies for reducing enteric methane generation.
Environmental Change
Environmental change, over the next 50-100 years, due to the warming effect of the accumulation of gases in the atmosphere, will clearly necessitate changes in resource allocation and utilization in the world and will directly affect all countries.
Although future climate change is being predicted from models, and these take into account a large number of variables, the useful information they generate is limited by the knowledge base and the unknowns. In fact, modeling is at present largely defining what is not known in terms of what will effect change in temperature, rainfall and sea levels, let alone sea currents, wind, sunshine hours, soil moisture, and the incidence of pests and diseases of man, animals and plants. There are, of course, a number of skeptics who claim that global warming is not going to occur.
There will be some beneficial effects through a warmer climate in some parts of the world, but on a global basis disadvantages will undoubtedly outweigh any advantages. The unknowns make it essential to quickly put in place technologies to slow green-house gas emissions and stabilize these in the atmosphere. The real point is that amelioration techniques put in place now have important associated benefits. In general, the strategies for decreasing greenhouse gas emissions mean reduction in fossil fuel utilization and general reduction of the release of pollutants into the atmosphere. Animal production strategies that target a decreased enteric methane release generally lead to improved production.
The approximate reductions in anthropogenic emissions required to stabilize atmospheric concentrations of gases to current levels are shown in Table 1.
The long time lag between gas released at the earth's surface mixing in the atmosphere and therefore, warming, together with the reluctance of governments to put into practice legislation to limit, in particular, carbon dioxide production from fossil fuels, suggests that there will be a rise in world environmental temperatures of 0.5 to 1deg.C in the next 25-50 years (see Gibbs et al. 1989).
In the last 30 years the expansion of crop and livestock production has more than doubled throughout the third world, although there are large differences between regions. Increases in human population (94 million in the last year), urbanization and improved income levels of the middle class have increased demand for food to such an extent that food surpluses are still rare.
Increases in animal production in the developing countries have mainly been a result of increasing animal numbers (see Jackson 1981). The lack of improvement in efficiency of ruminant production is well documented. In the developing countries low productivity of cattle is amplified by late age at puberty and long intercalving intervals in cows. It should be emphasized, however, that this is also a feature of ruminants fed low-quality forages in any country including Australia, New Zealand and South America.
New feeding strategies for animals fed on low quality forages (e.g. crop residues, tropical pastures etc.), coupled with better genotypes, improved management and disease control, particularly in India (see NDDB 1989) has changed this situation enormously (see Preston and Leng 1991). For example, large increases in milk production can be achieved in the tropics without the use of 'fossil-fuel-expensive' grain based concentrates, relying rather on by-products of agriculture (Leng 1991). These will be the only truly available feed resource for the large ruminant populations in these countries into the foreseeable future.
The Greenhouse Effect--A Simple Description
The greenhouse effect, or increasing world temperature, is due to the accumulation of gas in the atmosphere (Fig. 2). It is clearly ascribable to the major industrial countries, as some 50% of the increased retention of energy by the atmosphere is a result of the accumulation of carbon dioxide from combustion of fossil fuel. Industrialized countries presently use 70% of the world's oil production, and it has been much higher in the past (Fig. 3).
The other gases that contribute to increasing temperatures arise from a variety of activities; chloroflouro-carbons have been created by man, but will be phased out of production by legislation. Methane is an important component of greenhouse gases in the atmosphere, and is the one most associated with animal agriculture. Methane has a thermogenic effect some four to six times that of carbon dioxide.
Prior to the last two decades, world temperatures and composition of the atmosphere had changed little, but now there appears to be an ever-increasing rate of gas accumulation (Fig. 4). Undoubtedly contamination of the atmosphere with carbon dioxide, methane and the other greenhouse gases must be reduced or the future of the earth is threatened.
Methane concentrations in the world's atmosphere are rising rapidly, and although they contribute only 19% of the overall warming, methane is a major component. Methane arises largely from natural anaerobic ecosystems, rice paddies and fermentative digestion in ruminant animals (Fig. 5), whilst it is oxidized as an energy source by microorganisms in undisturbed soils (ref. Mosier et al. 1991). To stabilize its concentration in the atmosphere, methane production needs to be decreased by a mere 10-20%, compared with 80-85% reduction needed for the other gases (Table 1).
Animal Agriculture And The Greenhouse Effect
Animal agriculture contributes to accumulation of methane gas directly through production of methane in fermentative digestion in the rumen, and indirectly when faecal materials decompose anaerobically (see Safley et al. 1992).
Ruminants in 'natural' production systems are, in general, individually inefficient. Production increases have depended on increasing numbers, or increasing stocking rates without much increase in individual animal production on the more fertile grass lands.
There is a growing appreciation that efficiency of feed utilization per unit of production of meat, milk, work etc. can be improved considerably by simple technology inputs. If applied, this could have major implications for stabilizing global atmospheric methane concentrations.
The following discussion on how to reduce enteric methane production by ruminants will deal largely with feeding strategies that increase the efficiency of production and which eventually produce more product from fewer animals and less total feed.
'Environment-Friendly' development of livestock production systems demands that the increased production be met by increased efficiency of production and not through increased animal numbers. An increase in numbers of ruminants would put huge pressures on many resources including forests and land that might be afforested.
Methane production from ruminants
World ruminant population densities and estimated methane production rates by animals are shown in Table 2.
Global methane emissions
Methane is accumulating in the atmosphere at a rate of 1% per annum, and it contributes about 19% to global warming.
Ruminant animals produce a relatively small proportion (i.e. 15-20%) of the total global emissions. However, the domestic ruminants represent one of the few sources that can be manipulated. They are, in addition, an attractive target as reduction of methane is usually associated with improved productivity. It is estimated that of the enteric sources of methane, beef and draught animals contribute 50%, dairy cows 19% and only 9% is from sheep (Crutzen et al. 1986).
In general, growth rates, milk production and reproductive rates in these systems are low, compared with the genetic potential (e.g. liveweight gain is about 10% and rarely exceeds 30% of an animal's potential in a feed year).
In these systems cattle grow to maturity or slaughter weight over 3-5 years, cows produce a calf at 4-5 years, and on average every 2 years thereafter. Milk production on these feeding systems is often below 1000 L/lactation. Cows are often kept largely to produce draught oxen (e.g. in India 20 million cows are kept to provide replacements to the 80 million draught herd), and in some specialized systems they are kept only for the production of dung (which is valued as a fuel) and a number of other purposes (e.g. as a bank, for recreation and for religious purposes). It is therefore often difficult to sustain an argument that increasing the efficiency of feed utilization is the primary mechanism for decreasing global methane generation, as it cannot be certain that numbers of livestock will not increase when these technologies are applied.
Methane Production From Ruminants Fed 'Poor Quality' Forages
Methane output relative to product output of ruminants depends on two factors:
Efficiency of Rumen Fermentation of 'Poor Quality' Forages
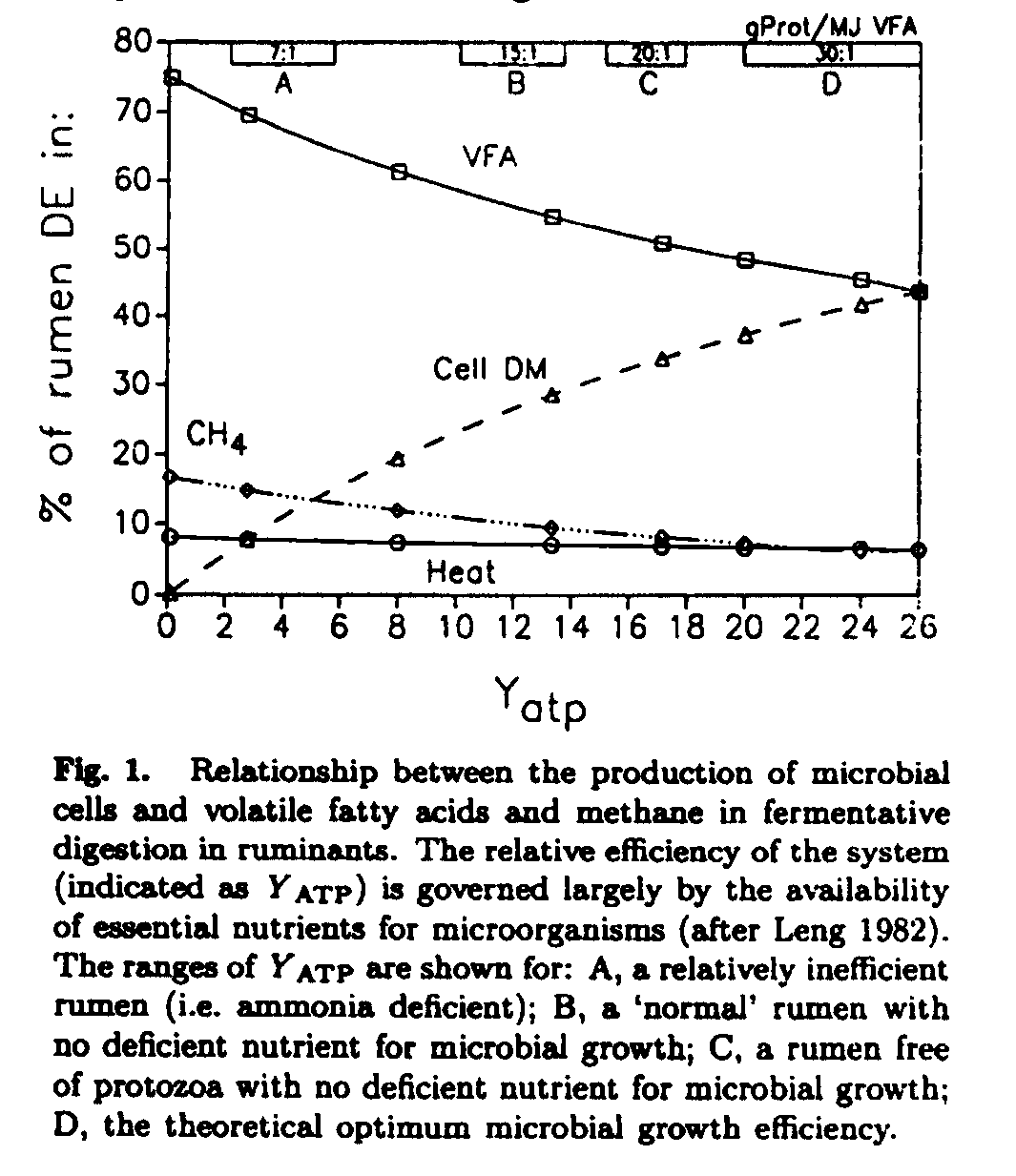
In cattle on a poor quality forage, a number of essential microbial nutrients may be deficient and, microbial growth efficiency in the rumen is low. In these conditions methane produced may represent 15-18% of the digestible energy, but correction of these deficiencies reduces this to as low as 7%. The relationships between products of fermentative digestion and the efficiency of the microbial ecosystem in the rumen is shown in Fig. 1.
Efficiency Of Feed Utilization By Ruminants Fed Crop Residues Or Other Low Protein Fibrous Feed
Research in the past 20 years has clearly illustrated that supplementation of cattle on low quality forage based diets increases productivity through increasing efficiency of feed utilization (Leng 1990b).
A mixture of nutrients as can be supplied for instance in a molasses urea multinutrient block lick ensures an efficient microbial digestion in the rumen. A small amount of protein meal that is directly available to the animal (i.e. bypass protein) stimulates both productivity and efficiency of feed utilization (the evidence and theory is discussed by Preston and Leng 1987).
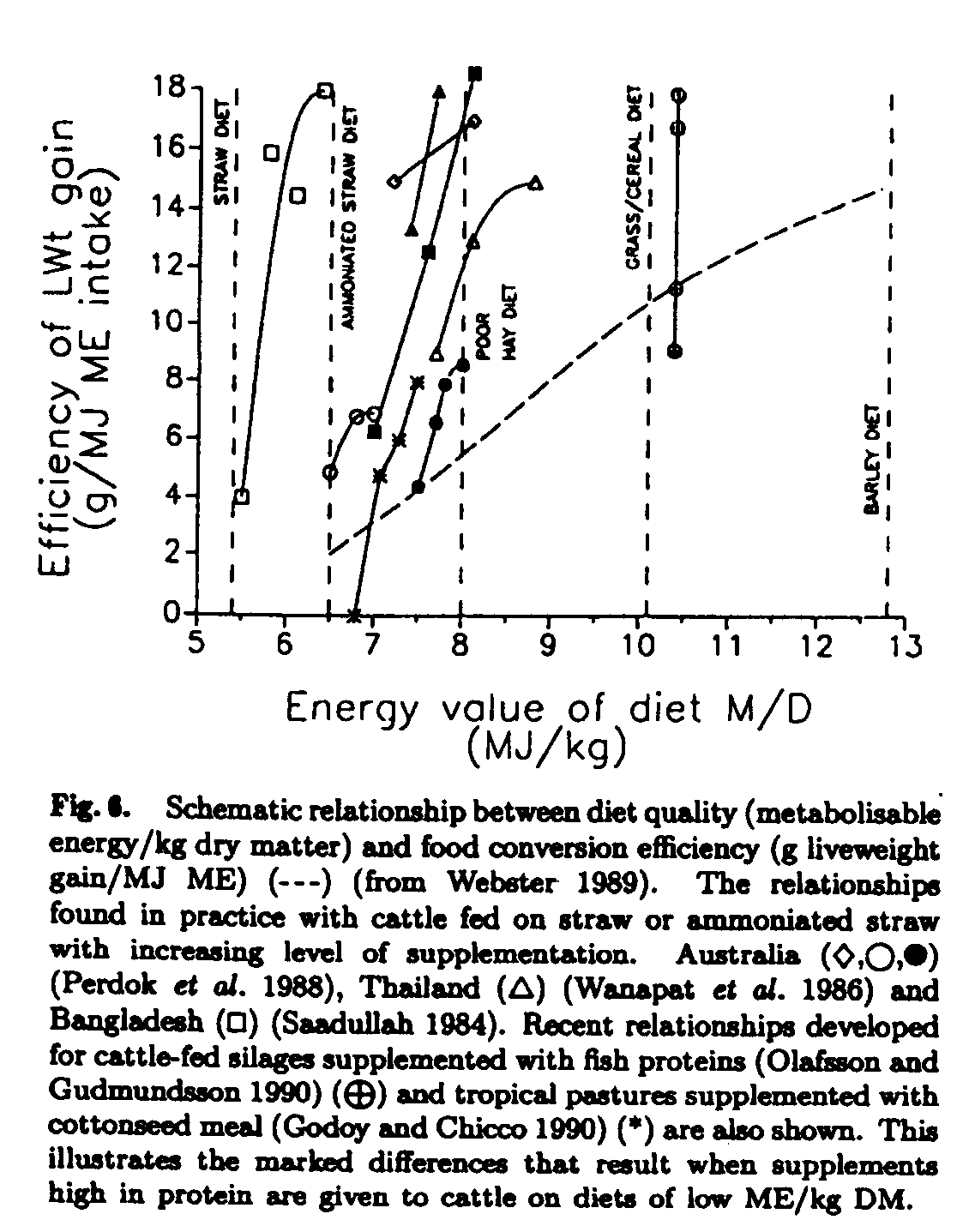
Traditional feeding standards are based on the metabolizable energy (ME) content of a feed. The general and theoretical relationship between ME/kg of feed and growth (g gain/unit of ME intake) are shown in Fig. 6. The results of a number of feeding trials with cattle on straw or low-quality pasture and silage-based diets supplemented with protein meals are shown in the same figure. The efficiency of growth relative to methane production assuming a relatively efficient rumen system calculated from the data in Fig. 6 is shown in Fig. 7 These data clearly show the massive reduction in methane production per unit of metabolizable energy intake and liveweight gain that is possible by using strategic supplements that accommodate the requirements of the rumen organisms and balance the absorbed nutrients to the animal's requirements.
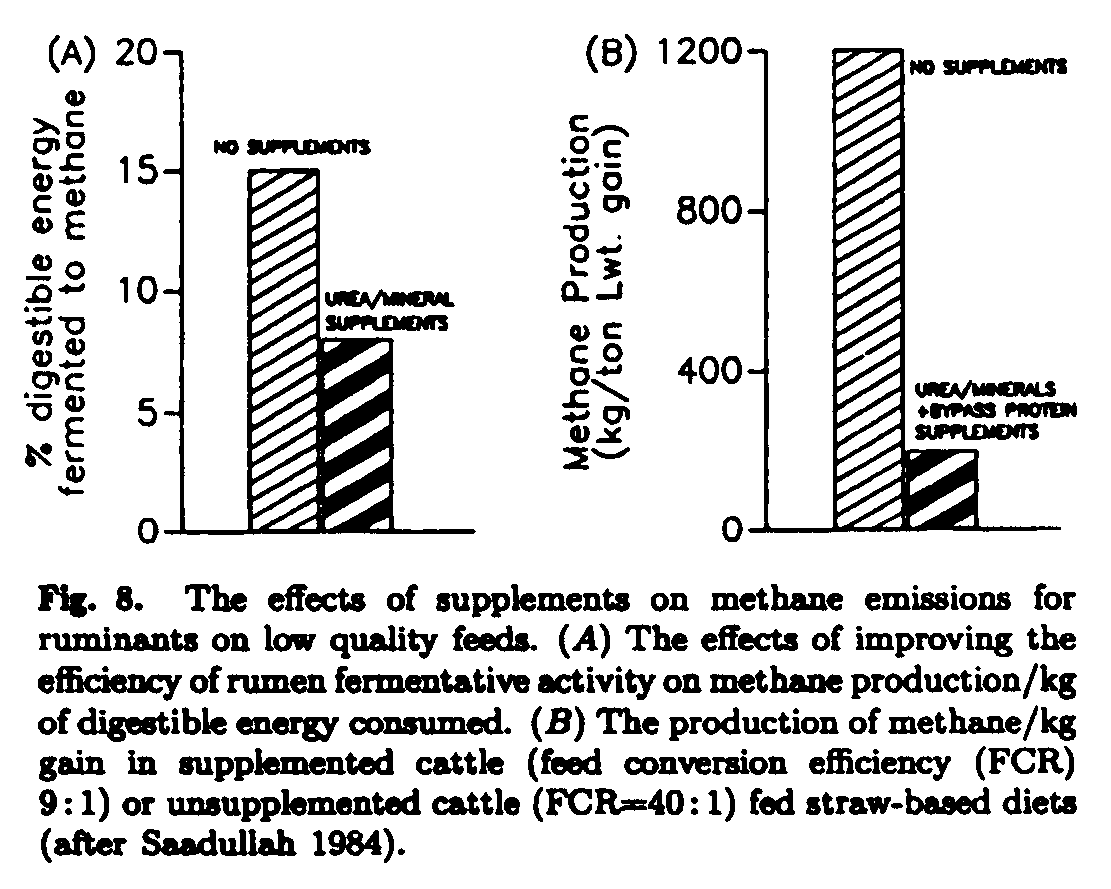
The data in Fig. 8 show the theoretical effects on methane production of balancing the rumen and the animal with protein supplements for experimental data with growing cattle fed straw in research carried out by Saadullah (1984). This indicates the massive potential reduction in methane per unit of liveweight gain that can result from introducing these supplementation strategies (Leng 1991).
Provision of molasses urea blocks to draught oxen which in general receive only straw in most developing countries will have a major effect on methane production, reducing it to perhaps half the present production rate. Preston (1991) has recently shown that this feeding strategy also improves work capacity of buffaloes.
Milk Production From Low Quality Forages
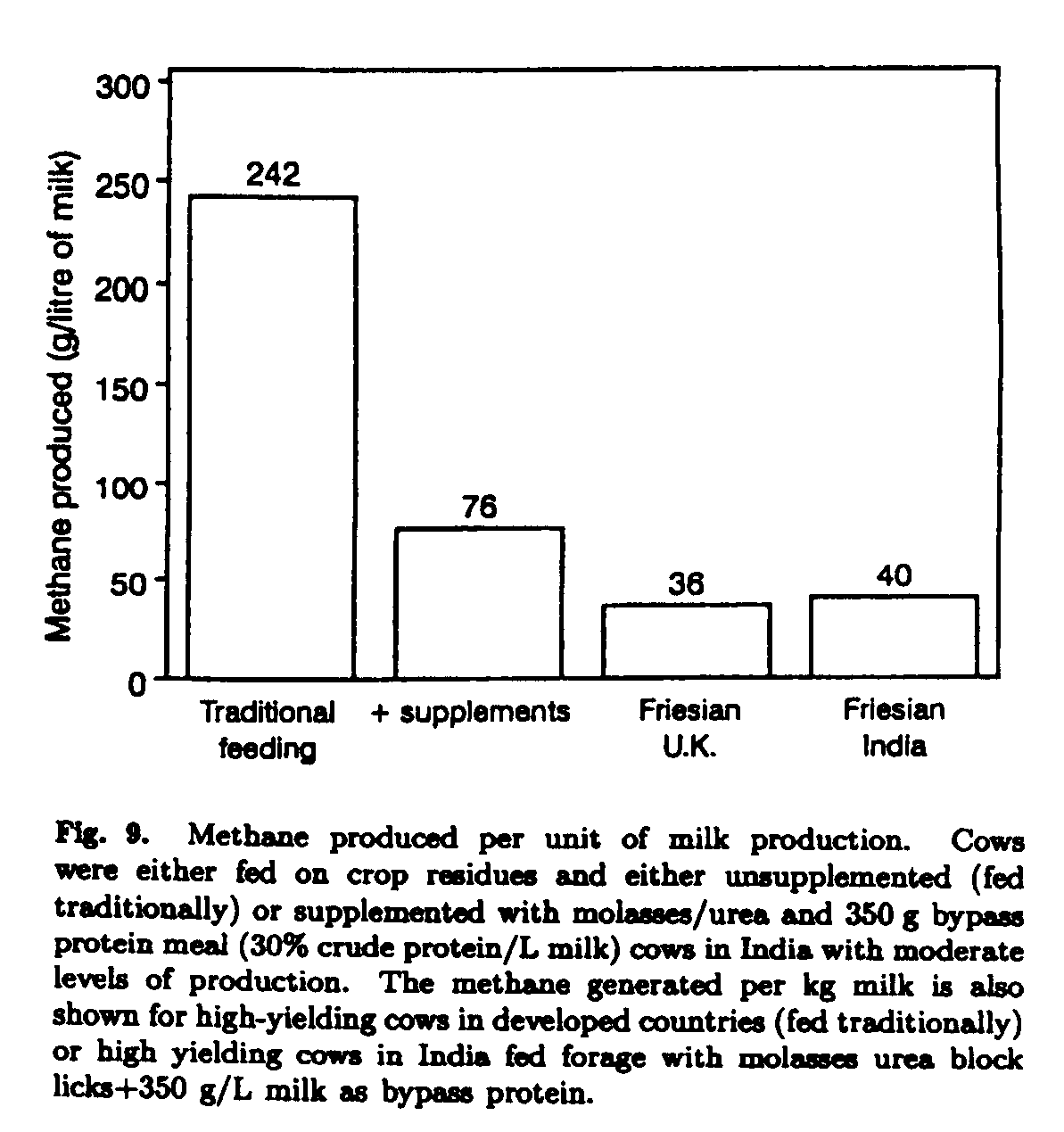
The same principles of supplementary feeding for growth have been found to stimulate milk production of dairy animals. The methane produced/unit milk yield under new feeding strategies that emphasize balancing the diet by supplementation with bypass protein or traditional feeding (i.e. concentrated food) are shown in Fig. 9. Calculations are made for local dairy cows, imported Friesians in India or Friesians under temperate country management. The data take into account lifetime methane production in relation to lifetime milk production. The calculations also include the effects of the supplementation on age at first calving, intercalving interval and improved milk yield of supplemented animals (Leng 1991).
Large ruminants produce some 15-20% of the global production of methane. Ruminants on low quality feeds possibly produce over 75% of the methane from the world's population of ruminants. Supplementation to improve digestive efficiency in, for example, draught animals could halve the methane production per unit of feed consumed. Together with supplementation to improve efficiency of feed utilization and increase product output, this may reduce methane production per unit of milk or meat by a factor of 4-6. Provided animal numbers decrease, as demand is met, the production of methane from the large populations of animals fed poor-quality forages could be reduced to below 50%, and perhaps even to as low as 25% of its present rate.
Overall Conclusions
Knowledge of quantitative nutrition provides a powerful tool to develop concepts to undertake a wide range of problem-oriented research. These vary from metabolic diseases of ruminants through to amelioration of enteric methane emissions from ruminants.
In a still rapidly expanding world, food production will need to be given some major priority in the future, and the problems that will arise will be solved only by researchers with a breadth and depth of understanding of scientific principles, a concept promulgated by Frank Annison from those early days in Armidale to the present time.
Baldwin, R. L. (1970). Energy metabolism in anaerobes. American Journal of Clinical Nutrition 23, 1508-13.
Black, J. L., Gill, M., Beever, D. E., Thornley, J. H. M., and Oldham, J. D. (1987a). Simulation of the metabolism of absorbed energy-yielding nutrients in young sheep. I. Efficiency of utilization of acetate. Journal of Nutrition 117, 105-15.
Black, J. L., Gill, M., Beever, D. E., Thornley, J. H. M., and Oldham, J. D. (1987b). Simulation of the metabolism of absorbed energy-yielding nutrients in young sheep. II. Efficiency of utilization of lipid. Journal of Nutrition 117, 116-28.
Bolle, H. J., Seiler, W., and Bolin, B. (1986). Other greenhouse gases and aerosols; assessing their role for atmospheric radiative transfer. In 'The Greenhouse Effect, Climatic Change and Ecosystems'. (Eds B. Bolin, B. R. Doos, B. Warrick and D. Jager.) pp. 157-203. (John Wiley and Sons: New York.)
Coleman, G. S. (1975). Interrelationship between rumen ciliate protozoa and bacteria. In 'Digestion and Metabolism in the Ruminant'. (Eds I. W. McDonald and A. C. I. Warner.) pp. 149-64. (U.N.E. Publishing Unit: Armidale, N.S.W.)
Crutzen, P. J., Aselmann, I., and Seiler, W. (1986). Methane production by domestic animals, wild ruminants, other herbivorous fauna, and humans. Tellus 38B, 271-84.
Czerkawski (1986). 'An Introduction to Rumen Studies.' p. 119 (Pergamon Press: Oxford.)
Fillery, I. R. (1991). Legume-fixed N and grazing animals. Important contributions to greenhouse gas nitrous oxide? Climate Change Newsletter 3(2), 1-2. (Bureau of Rural Resources: Canberra, A.C.T.)
Fraser, D. L., Poppi, D. P., Fraser, T., and Sykes, A. R (1991). Protein or amino acid supplementation of grazing iambs. In 'Recent Advances in Animal Nutrition in Australia'. (Ed D. J. Farrell.) p. 7A.(Department of Biochemistry, Microbiology and Nutrition University of New England: Armidale, N.S.W.)
Godoy, S., and Chicco, C. (1990). Annual Report. (FONIAP, Maracay: Venezuela.)
Gibbs, M. J., Lewis, L., and Hoffman, J. S. (1989). 'Reducing Methane Emissions from Livestock: Opportunities and Issues.' p. 284 (U.S. Environmental Protection Agency: Washington, D.C.)
Habib, G., Basit Ali Shah, S., Wahiduilah, Jabbar, G., and Ghufranullah (1991). The importance of urea-molasses blocks and bypass protein in animal production: the situation in Pakistan. In 'Isotope and Related Techniques in Animal Production and Health'. pp. 133-44. (IAEA: Vienna.)
Hendratno, N., Nolan, J. V., and Leng, R. A. (1991). The importance of urea-molasses multi nutrient blocks for ruminant production in Indonesia. In 'Isotope and Related Techniques in Animal Production and Health'. pp. 157-70. (IAEA: Vienna.)
Hennessy, D. W., Kempton, T. J., and Williamson, P. J. (1989). Copra meal as a supplement to cattle offered a low quality native pasture hay. Asian-Australian Journal of Animal Science 2, 77-84.
Hennessy, D. W., and Williamson, P. J. (1990). Feed intake and liveweight of cattle on subtropical native pasture hays. II. The effect of urea and maize flour, or protected casein. Australian Journal of Agricultural Research 41, 1179-85.
Hespell, R. B. (1979). Efficiency of growth by ruminal bacteria. Federation Proceedings 38, 2707-12.
Hungate, R. E. (1966). 'The Rumen and its Microbes.' p. 533. (Academic Press: New York.)
Jackson, M. G. (1981). A new livestock development strategy for India. World Animal Review 37, 2-8.
Klieve, A. V., Hudman, J. F., and Bauchop, T. (1989). Inducible bacteriaphages from ruminal bacteria. Applied and Environmental Microbiology 55, 1630-4.
Lashof, D. A., and Tirpak, D. A. (1990). 'Policy Options for Stabilising Global Climate, Report to Congress.' (United States Environmental Protection Agency: Washington, D.C.)
Leng, R A. (1982). Modification of rumen fermentation. In 'Nutritional Limits to Animal Production from Pastures'. (Ed. J.B. Hacker.) pp. 427-53. (CAB: Farnham Royal, U.K.)
Leng, R. A. (199Oa). Factors affecting the utilization of 'poor quality' forages by ruminants particularly under tropical conditions. Nutrition Research Reviews 3, 277-303.
Leng, R. A. (199Ob). Contribution of methane from ruminants to global methane production and some strategies for reducing emission from ruminants. In 'Rural Industries: Workshop on Climate Change'. pp. 31-40. (Bureau of Rural Resources: Canberra.)
Leng, R. A. (1991). 'Improving Ruminant Production and Reducing Emissions from Ruminants by Strategic Supplementation.' p. 105. (United States Environmental Protection Agency, Office of Air and Radiation: Washington, D.C.)
Leng, R. A., and Annison, E. F. (1963). Metabolism of acetate, propionate and butyrate by sheep-liver slices. Biochemical Journal 86, 319-27.
Leng, R. A., and Annison, E. F. (1964). The metabolism of D(-) B-hydroxybutyrate in sheep. Biochemical Journal 90, 464-9.
Leng, R. A., and Kunju, P. J. G. (1988). 'A New Approach on Protein Nutrition for Ruminants.' p. 22. (NDDB: Anand, India.)
Leng, R. A., and Nolan, J. V. (1984). Nitrogen metabolism in the rumen. Journal of Dairy Science 67 (5), 1072-89.
Loosli, J. K., and McDonald, I. W. (1968). 'Nonprotein Nitrogen in the Nutrition of Ruminants.' p. 94. (FAO: Rome.)
McRae, J. C. (1976). Utilization of the protein of green forage by ruminants at pasture. In 'Reviews in Rural Science. II. From Plant to Animal Protein'. (Eds T. M. Sutherland, J. R. McWilliam, and R A. Leng.) pp. 93-9. (University of New England Printing Unit: Armidale, N.S.W.)
Maang W. J., Chang, M. B., Yun, H. S., and Choi, I. (1989). Dilution rates on the efficiency of rumen microbial growth in continuous culture. Asian-Australian Journal of Animal Science 2, 477-80.
Mathison, G. W., and Milligan, L. P. (1971). Nitrogen metabolism in sheep. British Journal of Nutrition 25, 351-66.
Mosier, A., Schimel, D., Valentine, D., Bronson, K., and Parton, W. (1991). Methane and nitrous oxide fluxes in native, fertilized and cultivated grasslands. Nature (Lond.) 350, 330-2.
N.D.D.B. (1989). 'Annual Report of the National Dairy Development Board.' (National Dairy Development Board: Anand, India.)
Nolan, J. V., and Leng, R. A. (1972). Dynamic aspects of ammonia and urea metabolism in sheep. British Journal of Nutrition 27, 177-94.
Nolan, J. V., and Stachiw, S. (1979). Fermentation and nitrogen dynamics in Merino sheep given a low-quality roughage diet. British Journal of Nutrition 42, 63-80.
Olafsson, B. L., and Gudmundsson, O. (1990). Utilization of fisheries byproducts as supplements fed with roughages to ruminants. In 'Development of Feeding Strategies for Improving Ruminant Productivity in Areas of Fluctuating Nutrient Supply through the Use of Nuclear and Related Techniques'. (FAO/IAEA: Vienna.)
0rskov, E. R. (1970). Nitrogen utilization by the young ruminant. In 'Proceedings of the Fourth Nutrition Conference for Feed Manufacturers'. (Eds H. Swan and D. Lewis.) pp. 20-35. (J. and A. Churchill: London.)
Perdok, H. B., Leng, R. A., Bird, S. H., Habib, G., and Van Houtert, M. (1988). Improving livestock production from straw-based diets. In 'Increasing Small Ruminant Productivity in Semi- arid Areas'. (Eds E. F. Thomson and F. S. Thomson.) pp. 81-91. (ICARDA: Syria.)
Perdok, H., and Leng, R. A. (1989). Rumen ammonia requirements for efficient digestion and intake of straw by cattle. In 'The Roles of Protozoa and Fungi in Ruminant Digestion'. (Eds J. V. Nolan, R A. Leng, and D. I. Demeyer.) pp. 291-3. (Penambul Books: Armidale, N.S.W.)
Preston, T. R., and Leng, R. A. (1987). 'Matching Ruminant Production Systems with Available Resources in the Tropics and Subtropics.' p. 245. (Penambul Books: Armidale, N.S.W.)
Preston, T. R, and Leng, R. A. (1991). Agricultural technology transfer, perspectives and case studies. In 'Conference Proceedings from Agricultural Technology: Current Policy Issues for the International Community and the World Bank, October 1991'. (Ed. J. Anderson.) (World Bank: Washington D.C.) (in press.)
Resagno, A. and Segre, G. (1966). 'Drug and Tracer Kenetics.' (Walthani Blaisedell.)
Saadullah, M. (1984). Studies on utilisation of rice straw by cattle. Ph.D. Thesis, Royal Veterinary College, Copenhagen.
Saadullah, M. (1991). The importance of urea-molasses blocks and bypass protein in animal production: the situation in Bangladesh. In 'Isotope and Related Techniques in Animal Production and Health'. pp. 145-56. (IAEA: Vienna.)
Safley, L. M. Jr, Casada, M. E., Woodbury, J. W., and Roos. K. F. (1992). Global Methane Emissions from Livestock and Poultry Manure.' (U.S. Environmental Protection Agency: Washington, D.C.)
Silva, D. A. T., Greenhalgh, J .F. D., and 0rskov, E. R. (1989). Influence of ammonia treatment and supplementation on the intake digestibility and weight gain of sheep and cattle on barley straw diets. Animal Production 48, 99-108.
Veira, D. M., Ivan, M., and Jui, P. Y. (1983). Rumen ciliate protozoa: effects on digestion in the stomach of sheep. Journal of Dairy Science 66, 1015-22.
Wanapat, M., Duangchan, S., Pongpairote, S., Anakewit, T., and Tongpanung, P. (1986). Effects of various levels of concentrate fed with urea-treated rice straw for purebred American Brahman yearling cattle. In 'Ruminant Feeding Systems Utilizing Fibrous Agricultural Residues'. Proceedings of the 5th Annual Workshop of the Australian-Asian Fibrous Agricultural Residues Research Network. (Ed. R. W. Dixon.) pp. 149-53. (IDP: Canberra)
Manuscript received 24 August 1992, accepted 23 November 1992
{kind=link}
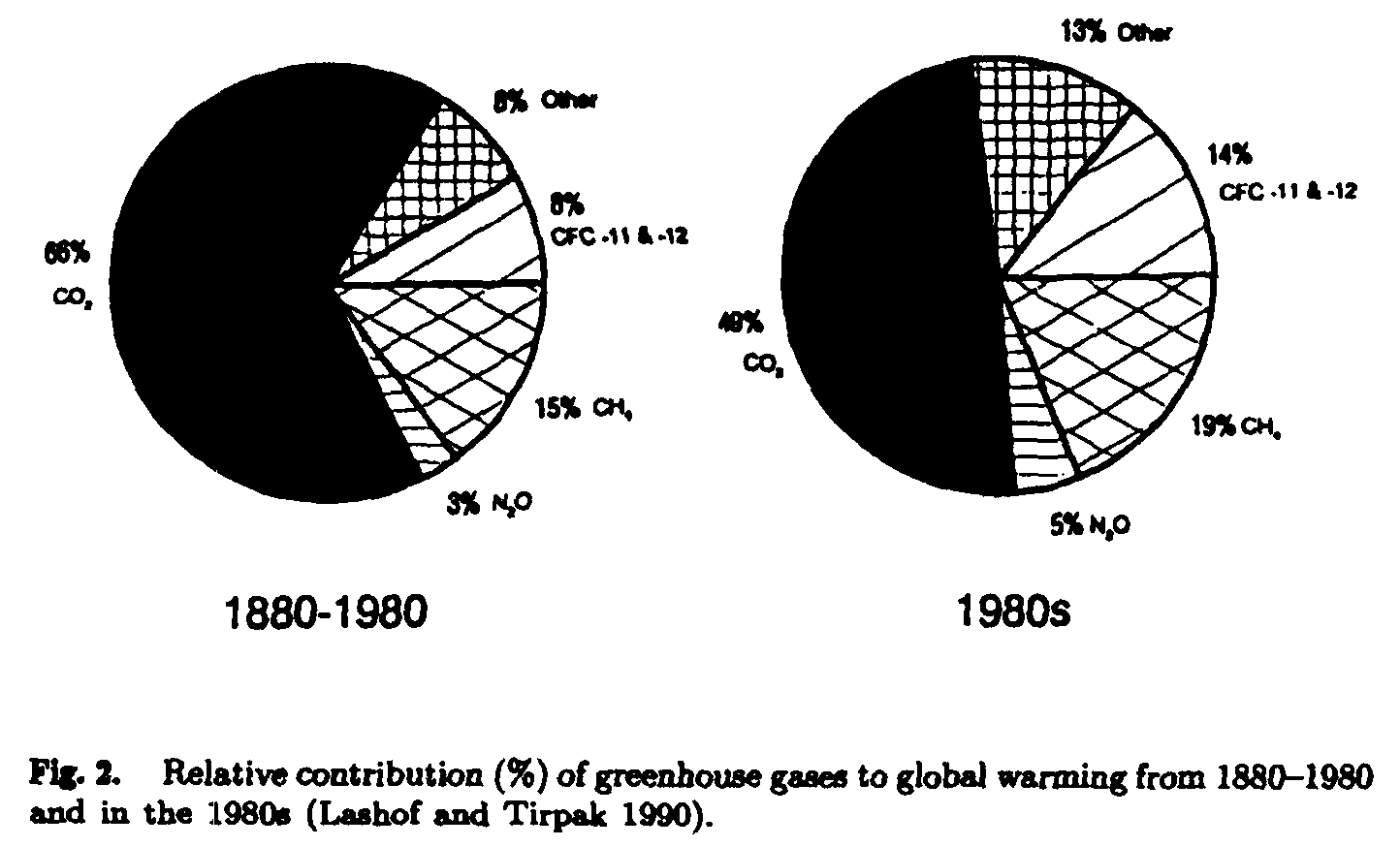{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}