 Reproduced, with permission, from:
Barnes, P. W., S. D. Flint, and M. M. Caldwell. 1990. Morphological responses of crop and weed species of different growth forms to ultraviolet-B radiation. American Journal of Botany 77: 1354-60.
Reproduced, with permission, from:
Barnes, P. W., S. D. Flint, and M. M. Caldwell. 1990. Morphological responses of crop and weed species of different growth forms to ultraviolet-B radiation. American Journal of Botany 77: 1354-60.
Department of Range Science and The Ecology Center, Utah State University,
Logan, Utah 84322-5230
The influence of ultraviolet-B (UV-B) radiation (280-320 nanometers) on the morphology of 12 common dicot and monocot crop or weed species was examined to determine whether any common responses could be found that might, in turn, be useful in predicting possible changes in competitive balance under solar UV-B enhancement. Under glasshouse conditions, UV-B exposure (simulating a 20% reduction in stratospheric ozone at Logan, Utah) was found to reduce leaf blade and internode lengths and increase leaf and axillary shoot production in several species. Overall, the directions of these trends were similar in the majority of species that exhibited a significant response. These morphological changes occurred without any significant reduction in total shoot dry matter production. There was no clear distinction in the response of crops and weeds, though monocots were found to be generally more responsive than dicots. Previous work in dense canopies has shown that the photomorphogenetic effects of UV-B alter leaf placement and thereby influence competition for light. Our results suggest that, under these conditions, changes in competitive balance resulting from increased UV-B might be expected more frequently when monocots are involved in mixtures, rather than mixtures of only dicots.
Recent evidence of a general, global decline of stratospheric ozone (Kerr, 1988; NASA, 1988) has heightened concern about possible ecological consequences of increases in solar ultraviolet-B (UV-B, 280-320 nm) radiation resulting from ozone depletion (Caldwell, Teramura, and Tevini, 1989). Although increased solar UV-B might lead to reduced photosynthesis and declines in production of some crop species (Teramura, 1983), the indirect effects of increased UV-B are considered by some to be of greater potential consequence (Caldwell, Teramura, and Tevini, 1989). Alteration in the balance of competition between plant species is an example of such indirect effects. Indeed, exposure to UV-B radiation, either at ambier (Bogenrieder and Klein, 1982) or enhance levels (Fox and Caldwell, 1978; Gold and Caldwell, 1983; Barnes et al., 1988), has been shown to alter competitive balance in many pairs of competing species. For most of these species, there was no effect of increased UV-B on yields when in monoculture.
Previous field studies with wheat (Triticum aestivum) and a weedy competitor, wild oat (Avena fatua), have shown that UV-B-induced shifts in competitive balance were associated with differential effects of the UV-B on shoot morphology which led to shifts in the relative positioning of leaf area for the two species in the mixed canopies (Barnes et al., 1988). The major morphological changes were altered leaf blade length and internode length as gauged by leaf insertion height in the canopy. These changes in canopy structure were computed to be sufficient to alter light interception and canopy photosynthesis for these species (Ryel et al., 1990). There were no direct effects of the increased UV-B on individual leaf photosynthesis (Beyschlag et al., 1988). Thus, we suggested that UV-B enhancement alters competitive balance by inducing changes in plant morphology and leaf placement in the canopy, which in turn changes the competition for light between the two species. The photomorphogenetic changes themselves may not necessarily be considered as damaging, and there is precedent for such alterations to be mediated by a specific UV-B photoreceptor (Wellmann, 1983; Steinmetz and Wellmann, 1986).
Whether this proposed mechanism of competitive change has general application to other species mixtures is not known. Of immediate interest in evaluating the economic impact of ozone reduction is the balance of competition between crop and weed species. In the present study, we examined the effect of enhanced UV-B irradiation on the morphology of a variety of common crop and weed species, both monocots and dicots. We specifically wished to determine whether any patterns of response in common could be found among these categories of species that might be useful in predicting possible changes in competitive balance under solar UV-B enhancement. Morphological changes with direct bearing on canopy architecture were of particular interest.
Twelve crop or weed species (Table 1) were grown in pure 'stands' in a glasshouse and exposed to supplemental UV-B radiation or control conditions with no UV-B radiation. Varieties of crop species in common agronomic use, and weed species from seed collected from agricultural fields near Logan, Utah, were used. Plants were grown in a 3:1 (v:v) mixture of organic potting medium and sand in individual containers (0.15-1 volume). Rows of buffer plants were used to avoid edge effects. The containers were arranged in racks with regular interplant spacings of 4.5 cm and a plant density of 494 m[-2], which was sufficiently high to result in a closed canopy. Since competition for light between crop and weed species often occurs in essentially closed canopies, we felt that shading within the canopy was pertinent for these experiments. Furthermore, the photomorphogenetic responses being observed in these studies often involve the participation of several photosystems such as the phytochrome, cryptochrome, and the UV-B receptor (Cosgrove, 1986). Individual plants were kept well watered and fertilized (Hoagland's nutrient solution) throughout the experiment. Natural sunlight was supplemented by 1,000-W high pressure sodium vapor lamps for 12 hr day[-1].
Because of limited space under the UV-lamp systems, only four species could be grown and exposed to UV-B radiation concurrently. Thus, three exposure periods were used in this study; the species and corresponding light and temperature conditions for these exposure periods are shown in Table 2. Photon flux density (PFD; 400-700 nm) was measured at the top of the plant canopy with gallium arsenide phosphide photodiodes (diffusion type, Hamamatsu, G1118, Middlesex, NJ) calibrated against a quantum sensor (LiCor, Lincoln, NE) under natural sunlight. Air temperature was measured with copper-constantan thermocouples placed at canopy height. All sensors were interrogated at 5-min intervals by a small data logger (Model 21X, Campbell Scientific, Logan, UT), and hourly averages were recorded.
Ultraviolet radiation was supplied by fluorescent lamps (UV-B 313, Q-Panel, Cleveland, OH) enclosed by either 0.13-mm-thick cellulose acetate film (diacetate type) (UV-B treatment) or 0.13-mm-thick clear polyester (optically equivalent to Mylar D) (control; no UV-B) as described by Barnes et al. (1988). The estimated effective UV-B irradiance was 9.6 kJ m[-2] d[-1], which corresponds to a 20% depletion in the ozone column on 1 June for clear skies at Logan, UT (1,460 m elevation, 41.5'N latitude) based on the model of Green, Cross, and Smith (1980) and using the generalized plant action spectrum of Caldwell (1971) as the weighting function normalized at 300 nm. Ultraviolet irradiance was measured with a double-monochromator spectroradiometer (Optronic Model 742, Orlando, FL). This instrument was calibrated for absolute responsivity against a l,000-W tungsten-halogen standard lamp (Optronic Model 200 U, Orlando, F L) which was periodically calibrated against a l,000-W tungsten-halogen standard lamp from the National Institute of Standards and Technology (formerly National Bureau of Standards). Wavelength accuracy was checked before this calibration, and again when the instrument was moved to the glasshouse, by scanning a low-pressure mercury lamp that has a known discharge at distinct, well-separated emission lines.
Because the species differed in shoot height, containers of the shorter species were elevated as the experiment progressed so that UV-B irradiance at the top of the canopy was similar for all species. Since there was only one treatment and one control lamp bank, confoundment with position in the glasshouse could occur. To offset this, the containers were placed on mobile cans that were rotated 180deg. daily and switched between the two UV-lamp systems every 3 days when the UV-B transparent and UV-B-absorbing filters were replaced and alternated between the two lamp banks.
Irradiation treatments were initiated as soon as uniform cohorts of seedlings were available and continued for 20 to 44 days (Table 2) depending on size attained. At the time of harvest, the following measurements were taken on the main (primary) shoots of most species: maximum height, length and width of leaf blades, and the insertion height of leaves (measured as the height of the point of attachment of the leaf blade to the sheath for monocots or of the petiole to the stem for dicots). The number of dead and live leaves on the main shoot were counted; these included both fully expanded and emerging leaves for monocots and all leaves > 1 cm long for dicots. Main shoots were separated into leaf blade, petiole (dicots), sheath (monocots), and stem; reproductive tissue components were also considered for Setaria and Amaranthus. Daughter tillers of monocots, or axillary branch shoots of dicots, were separated from the main shoot tissue except for Triticum cylindricum, where there was no clear distinction between main and daughter tillers. Samples were oven dried (70 C) for 48 hr and weighed. Main shoot foliage area was determined using a leaf area meter (LICOR Model 3100, Lincoln, NE) for the dicots (except Kochia). For the monocots and Kochia, main shoot foliage area was estimated using the product of the weight of live leaves and a leaf area: dry weight ratio measured on a subset of plants.
Mean comparisons were made with Student's t-tests on weight, measurement, and enumeration data. Where weights were convened to fractional values, data were subjected to the arcsin-square-root transformation prior to analysis (Zar, 1984). Where leaf area was calculated as the product of the leaf weight and the area-to-weight ratio (which were measured on different sets of plants), an unbiased estimate for the variance of the products was used (Goodman, 1960). To facilitate a comparison of responses among species, we have expressed the morphological data as the deviation of UVB-treated plants from values of the control plants. Statistical tests, however, were conducted only on the measured, quantitative data.
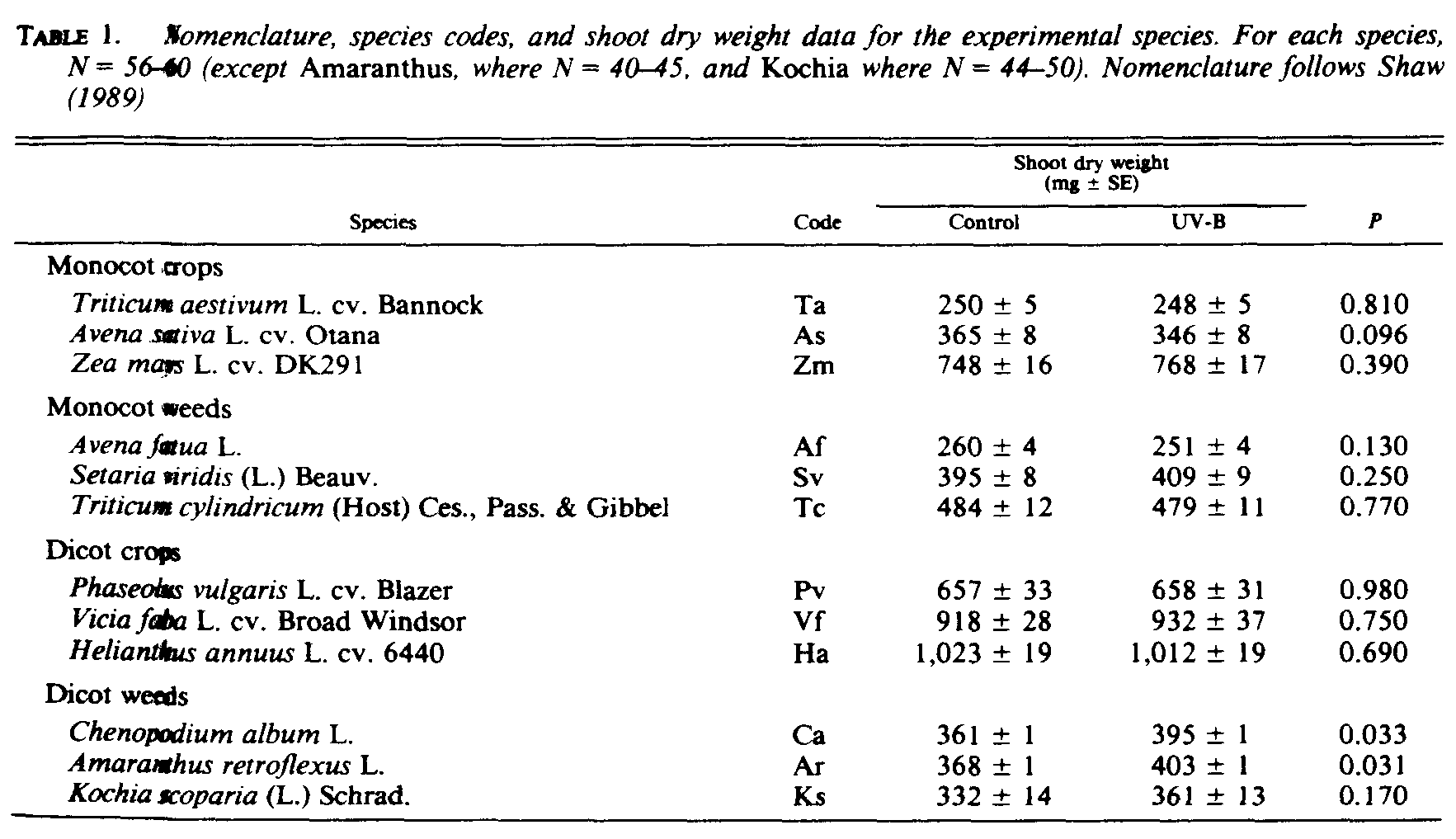
Total shoot dry weight was not significantly (P < 0.05) reduced by the UV-B treatment for any species; however, there were significant increases for two of the dicot weedy species, Chenopodium and Amaranthus (Table 1).
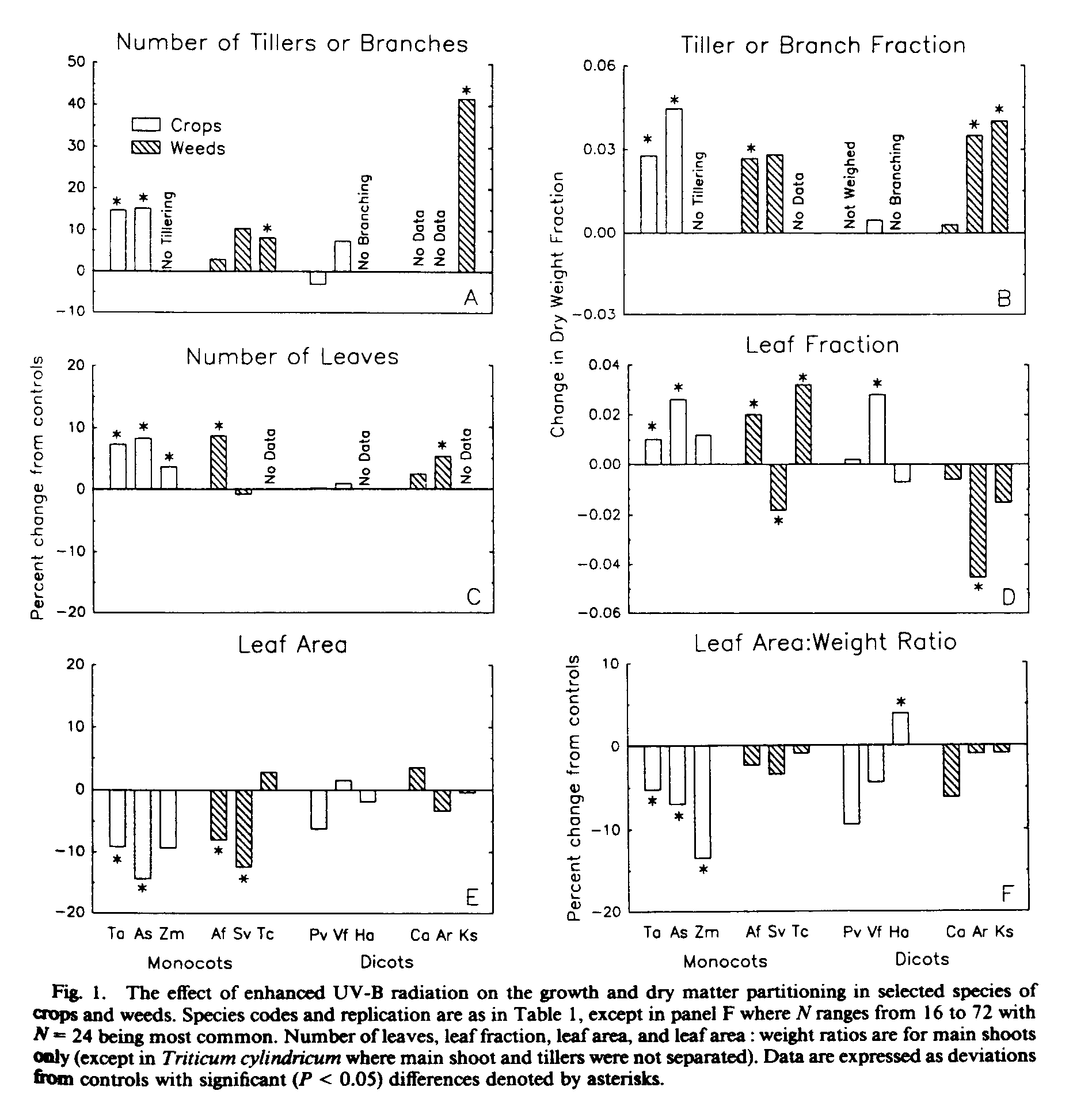
Although shoot dry weight was not reduced by the UV-B treatment, there were definite effects of the UV-B on plant morphology and dry matter partitioning. For several of the monocot species, exposure to UV-B resulted in an increased number of daughter tillers (Fig. 1a) and a greater shoot dry weight fraction of the daughter tillers (Fig. 1b). A similar response was not seen in the dicot species except for Kochia, in which case axillary shoot production and branch fraction were stimulated by UV-B, and for Amaranthus where branch fraction was increased (Fig. 1a, b). Four of the species not stimulated by the UV-B treatment, Vicia, Chenopodium, Zea, and Helianthus, produced very little (2% or less) or no detectable axillary shoot growth even under control conditions.
At the time of harvest, most monocot species had more leaves on the main shoot under UV-B treatment than in the controls (Fig. lc). For the dicots, a stimulation of leaf production by UV-B was only found for Amaranthus. For most monocots, the increase in the number of leaves coupled with a tendency for these leaves to be thicker (Fig. 1f) resulted in an increase in the leaf blade weight fraction of main shoots (Fig. 1d). The two species showing significant decreases in allocation to main shoot leaf fraction (Setaria and Amaranthus) had significant increases in main shoot reproductive tissues (data not shown). Leaf area of the main shoot was significantly reduced by UV-B treatment in some monocots, and was generally not affected in the dicots (Fig. 1e).
The UV-B treatment was found to generally suppress main shoot height, internode length (measured as leaf insertion height), and leaf length, though there was considerable variation among species in the magnitude of response (Fig. 2). Avena sativa and A. fatua showed large reductions in height and leaf length while Zea and Helianthus were rather insensitive. By comparison, Amaranthus and Kochia showed significant increases in shoot height under UV-B treatment. Significant reductions in leaf blade length occurred for several monocots, but the dicots did not exhibit a significant decrease of either leaf lamina (Fig.2c) or petiole (data not shown).
UV-B radiation is known to be detrimental to plants and has been shown to cause inhibition of photosynthesis and growth for several species, at least under laboratory or glasshouse conditions (e.g., Sisson and Caldwell, 1976; Kossuth and Biggs, 1981; Teramura and Murali, 1986); or using solar radiation in controlled environment chambers in the field (Tevini et al., in press). However, under field conditions the same general levels of supplemental UV-B seldom result in detectable reductions of photosynthesis (Caldwell, Teramura, and Tevini, 1989). If photosynthesis is inhibited to an appreciable extent in laboratory experiments, reductions of growth also occur. In our studies, we have found that morphological alterations, including reduced growth of some plant parts (e.g., internodes) and stimulated growth of other parts (e.g., axillary shoots) can occur without any apparent inhibition of leaf carbon assimilation (Beyschlag et al., 1988) or total shoot biomass production (Table 1; Fig. 1; also see Bames et al., 1988). Reductions in shoot height, stem internode length, and leaf size (Fig. 2) have been observed in a variety of different plant species (Becwar, Moore, and Burke, 1982; Teramura, 1983; Latimer and Mitchell, 1987; Sullivan and Teramura, 1988). Stimulation of axillary shoot growth and leaf production by UV-B, which we have found in several species (Fig. 1), has occasionally been reported by other investigators (Biggs and Kossuth, 1978; Teramura 1980; Barnes et al., 1988; J. Sullivan, unpublished data) but has not previously been recognized as a general plant response to UV-B.
The morphological changes we observed may reflect very specific photomorphogenetic responses of plants to UV-B radiation (Wellmann, 1983; Steinmetz and Wellmann, 1986). A UV-B photoreceptor has been shown to interact with the phytochrome and blue light receptors (Mohr, 1986), two photomorphogenetic systems known to influence stem internode elongation and tillering (Casal, Sanchez, and Derigibus, 1987; Child and Smith, 1987, Warpeha and Kaufman, 1989). UV-B radiation has also been found to interact directly with the metabolism of plant growth regulators. such as indoleacetic acid (IAA) (Curry, Thimann, and Ray, 1956; Tevini et al., in press). Regardless of the mechanism by which UV-B radiation alters growth, it does appear that, in many plant species, morphology is a more sensitive indicator of realistic UV-B radiation exposure than either leaf photosynthesis or total biomass production, at least under conditions of relatively high PFD.
In dense canopies, even subtle effects on internode elongation and/or leaf size can lead to changes in light interception and canopy photosynthesis of competing species (Barnes et al., 1990; Ryel et al., 1990). These, in turn, can eventually result in shifts in competitive balance as mentioned at the outset of this paper. The field studies that have shown these changes of competitive balance extract a considerable toll of time and effort and are necessarily specific to the species and growing conditions employed. Thus, this glasshouse screening study was undertaken to determine if morphological changes induced by UV-B are evident in a variety of species and if the changes suggest that some types of plants may be more affected than others. We felt it important to grow the plants in high density, as in the field, so that plants experienced shading by neighboring plants as well as the imposed irradiation treatments. Earlier work with wheat and wild oat conducted with isolated and grouped plants in a glasshouse showed several differences in response to the UV-B treatments (Barnes et al., 1988).
Although this study is focused on shoot morphological changes that can mediate changes in light competition, the UV-B could conceivably affect other aspects of competition such as below-ground interference. Such phenomena certainly warrant attention in future work. However, our studies with wheat and wild oat mixtures indicated that the changes in shoot morphology leading to altered light interception were sufficient to account for the changes in competitive balance (Barnes et al., 1988; Ryel et al., 1990).
While a quantitative extrapolation from these responses in the glasshouse to competitive balance in the field is clearly inappropriate, some qualitative similarities in the responses of plants in glasshouse and field situations have been established (Barnes et al., 1988). For example, wheat and wild oat plants grown in dense mixed stands in the glasshouse with individual plant roots in separate containers responded in a similar manner to mixtures of these two species in field experiments--wheat shifted more leaf area into upper canopy layers than did wild oat under enhanced UV-B.
The results of the present screening study indicated that the morphological responses of crop and weedy species to UV-B radiation did not differ in a particular pattern. Monocots. however, did tend to be more morphologically responsive to the UV-B than dicots. These findings suggest the following working hypotheses: 1) Changes of competitive balance between crop and weed associations resulting from increased UV-B should not necessarily be expected to favor either crop or weed species in a consistent manner; 2) Changes in competitive balance resulting from increased UV-B may be expected more frequently when monocots are involved in mixtures, rather than mixtures of only dicots; and 3) Enhanced UV-B may not alter monoculture production of species since even if morphological changes induced by increased UV-B result in changing heights of canopy foliage, the total intercepted radiation will not change.
BARNES, P. W., W. BEYSCHLAG, R. RYEL, S. D. FLINT, AND M. M. CALDWELL. 1990. Plant competition for light analyzed with a multispecies canopy model III. Influence of canopy structure in mixtures and monocultures of wheat and wild oat. Oecologia 82: 560-566.
------, P. W. JORDAN, W. G. GOLD, S. D. FLINT, AND M. M. CALDWELL. 1988. Competition, morphology and canopy structure in wheat (Triticum aestivum L.) and wild oat (Avena fatua L.) exposed to enhanced ultraviolet-B radiation. Functional Ecology 2: 319-330.
BECWAR, M. R., F. D. MOORE III, AND M. J. BURKE. 1982. Effects of deletion and enhancement of ultraviolet-B (280-315 nm) radiation on plants grown at 3,000 m elevation. Journal of the American Society for Horticultural Science 107: 771-774.
BEYSCHLAG, W., P. W. BARNES, S. D. FLINT, AND M. M. CALDWELL. 1988. Enhanced UV-B irradiation has no effect on photosynthetic characteristics of wheat (Triticum aestivum L.) and wild oat (Avena fatua L.) under greenhouse and field conditions. Photosynthetica 22: 516-525.
BIGGS, R. H., AND S. V. KOSSUTH. 1978. UV-B biological and climate effects research. Terrestrial FY 1977. Impact of solar UV-B radiation on crop productivity. Final Report. U.S. Department of Agriculture/Environmental Protection Agency, Washington, DC.
BOGENRIEDER, A., AND R. KLEIN. 1982. Does solar UV influence the competitive relationship in higher plants? In J. Calkins [ed.], The role of solar ultraviolet in marine ecosystems, 641-649. Plenum Press, New York and London.
CALDWELL, M. M. 1971. Solar ultraviolet radiation and the growth and development of higher plants. In A. C. Giese [ed.], Photophysiology, 131-177. Academic Press, New York.
------, A. H. TERAMURA, AND M. TEVINI. 1989. The changing solar ultraviolet climate and the ecological consequences for higher plants. Trends in Ecology and Evolution 4: 363-367.
CASAL. J. J., R. A. SANCHEZ, AND V. A. DEREGILUS. 1987. Tillering responses of Lolium multiflorum plants to changes of red/far-red ratio typical of sparse canopies. Journal of Experimental Botany 38: 1432-1439.
CHILD, R., AND H. SMITH. 1987. Phytochrome action in light-grown mustard: kinetics, fluence-rate compensation and ecological significance. Planta 172: 219-229.
COSGROVE, D. J. 1986. Photomodulation of growth. In R. E. Kendrick and G. M. H. Kronenberg [eds.], Photomorphogenesis in plants, 341-366. Martinus Nijhoff, Dordrecht.
CURRY, G. M., K. V. THIMANN, AND P. M. RAY. 1956. The base curvature response of Avena seedlings to the ultraviolet. Physiologia Plantarum 9: 429-440.
FOX, F. M., AND M. M. CALDWELL. 1978. Competitive interaction in plant populations exposed to supplementary ultraviolet-B radiation. Oecologia 36: 173-190.
GOLD, W. G., AND M. M. CALDWELL. 1983. The effects of ultraviolet-B radiation on plant competition in terrestrial ecosystems. Physiologia Plantarum 58: 435-444.
GOODMAN, L.A. 1960. On the exact variance of products. American Statistical Association Journal 55: 708-713.
GREEN, A. E. S., K. R. CROSS, AND L. A. SMITH. 1980. Improved analytic characterization of ultraviolet skylight. Photochemistry and Photobiology 31: 59-65.
KERR, R. A. 1988. Stratospheric ozone is decreasing. Science 239: 1489-1491.
KOSSUTH, S. V., AND R. H. BIGGS. 1981. Ultraviolet-B radiation effects on early seedling growth of Pinaceae species. Canadian Journal of Forest Research 11: 243-248.
LATIMER, J. G.. AND C. A. MITCHELL. 1987. UV-B radiation and photosynthetic irradiance acclimate eggplant for outdoor exposure. HortScience 22: 426-429.
MOHR, H. 1986. Coaction between pigment systems. In R. E. Kendrick and G. H. M. Kronenberg [eds.], Photomorphogenesis in plants, 547-564. Martinus Nijhoff, Dordrecht.
NASA. 1988. Present state of knowledge of the upper atmosphere 1988: an assessment report. NASA Reference Publication 1208.
RYEL, R. J., P. W. BARNES, W. BEYSCHLAG, M. M. CALDWELL, AND S. D. FLINT. 1990. Plant competition for light analyzed with a multispecies canopy model. I. Model development and influence of enhanced UV-B conditions on photosynthesis in mixed wheat and wild oat canopies. Oecologia 82: 304-310.
SHAW, R. J. 1989. Vascular plants of northern Utah: an identification manual. Utah State University Press, Logan, UT.
SISSON, W. B., AND M. M. CALDWELL. 1976. Photosynthesis, dark respiration, and growth of Rumex patientia L. exposed to ultraviolet irradiance (288 to 315 nanometers) simulating a reduced atmospheric ozone column. Plant Physiology 58: 563-568.
STEINMETZ V., AND E. WELLMANN. 1986. The role of solar UV-B in growth regulation of cress (Lepidium sativum L.) seedlings. Photochemistry and Photobiology 43: 189-193.
SULLIVAN, J. H., AND A. H. TERAMURA. 1988. Effects of ultraviolet-B irradiation on seedling growth in the Pinaceae. American Journal of Botany 75: 225-230.
TERAMURA, A. H. 1980. Effects of ultraviolet-B irradiances on soybean. I. Importance of photosynthetically active radiation in evaluating ultraviolet-B irradiance effects on soybean and wheat growth. Physiologia Plantarum 48: 333-339.
------. 1983. Effects of ultraviolet-B radiation on the growth and yield of crop plants. Physiologia Plantarum 58: 415-427.
------, AND N. S. MURALI. 1986. Intraspecific differences in growth and yield of soybean exposed to ultraviolet-B radiation under greenhouse and field conditions. Environmental and Experimental Botany 26: 89-95.
TEVINI, M., U. MARK, G. FIESER, AND M. SAILE. In press. Effects of enhanced solar UV-B radiation on growth and function of selected crop plant seedlings. In E. Riklis [ed.], Proceedings of the 10th International Congress of Photobiology, Jerusalem 1988.
WARPEHA, K. M. F., AND L. S. KAUFMAN. 1989. Bluelight regulation of epicotyl elongation in Pisum sativum. Plant Physiology 89: 544-548.
WELLMANN, E. 1983. UV radiation in photomorphogenesis. In W. Shropshire and H. Mohr [eds.], Encyclopedia of plant physiology, vol. 16B (new series), Photomorphogenesis, 745-756. Springer-Verlag, Berlin.
ZAR. J. H. 1984. Biostatistical analysis. Prentice Hall, Englewood Cliffs, NJ.
{kind=link}
{kind=link}
{kind=link}
{kind=link}