 Reproduced, with permission, from:
Herndl, G. J., G. Muller-Niklas, and J. Frick. 1993. Major role of ultraviolet-B in controlling bacterioplankton growth in the surface layer of the ocean. Nature 361: 717-19.
Reproduced, with permission, from:
Herndl, G. J., G. Muller-Niklas, and J. Frick. 1993. Major role of ultraviolet-B in controlling bacterioplankton growth in the surface layer of the ocean. Nature 361: 717-19.
Gerhard J. Herndl, Gerald Muller-Niklas & Jurgen Frick
Institute of Zoology, University of Vienna, Althanstrasse 14,
A-1090 Vienna, Austria
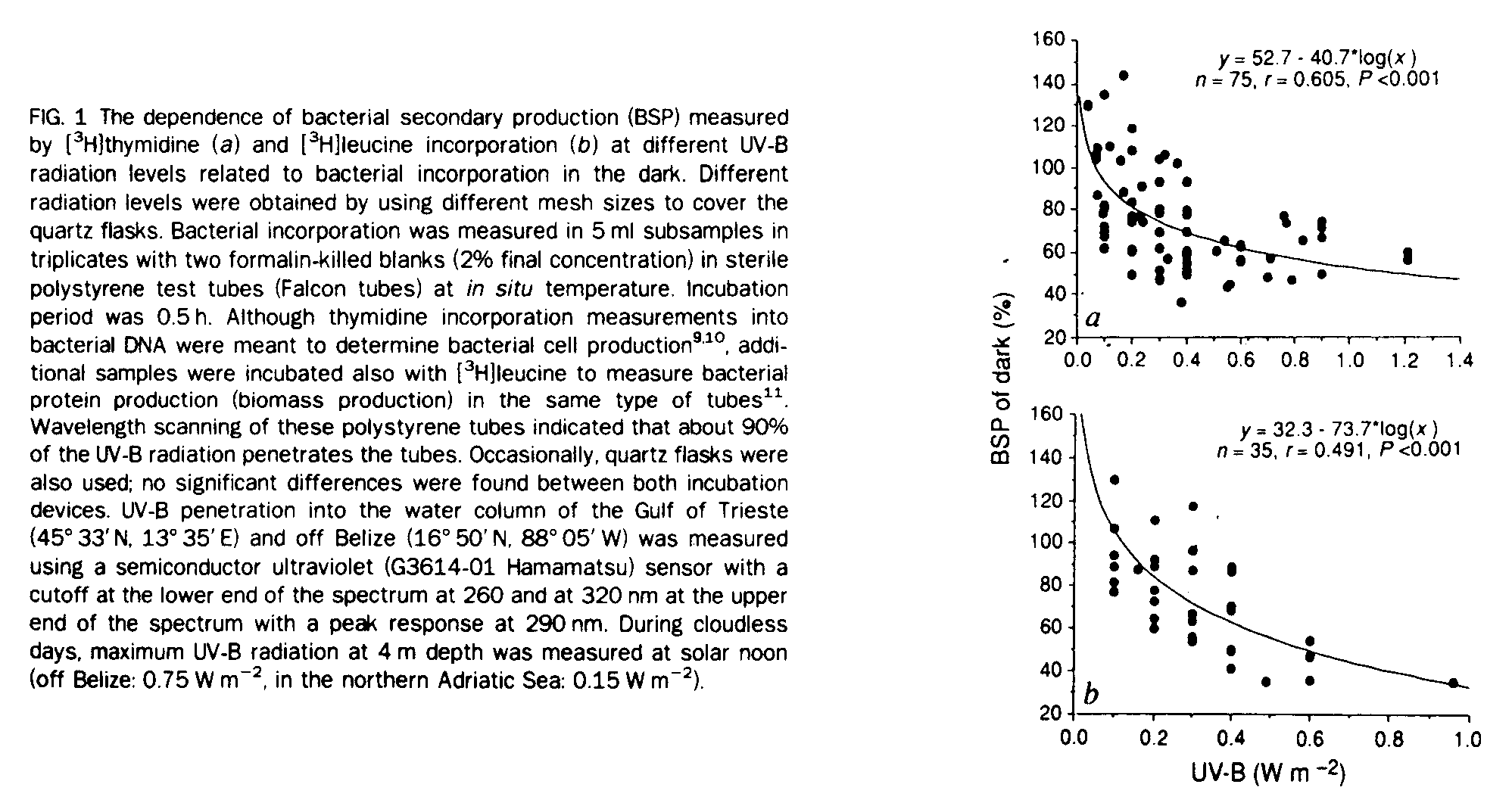
In evaluating the possible effects of natural solar radiation, we have to distinguish possible effects of short- and long-term exposure on the bacterial community. In a mixed water column, short-term exposure should predominate. We determined the influence of short-term (30 min) solar radiation on thymidine and leucine incorporation into bacterioplankton of the surface layer of the sea by incubating surface water at different UV-B radiation levels. Surface water samples were collected with acid-cleaned Niskin bottles in the northern Adriatic Sea from depths ranging from 0.5 m to 2 m. Bacterial cell production as well as bacterial protein production was affected by UV-B radiation typically for surface layers (Fig. 1). Compared with bacterial activity in the dark, a 50% reduction in thymidine incorporation was observed at UV-B radiation levels of 1.3 W m-2, whereas a 50% reduction in leucine incorporation was detectable at a radiation of about 0.6 W m-2 (Fig. 1). Thus we conclude that, at least during short-term exposure, bacterial biomass production is more affected than cell division and hence cell production. The higher bacterial activity in the low UV-B radiation range (<0.2 W m-2) compared to the dark activity might be caused by the release of easily metabolizable substances from the photosynthetic activity of phytoplankton.
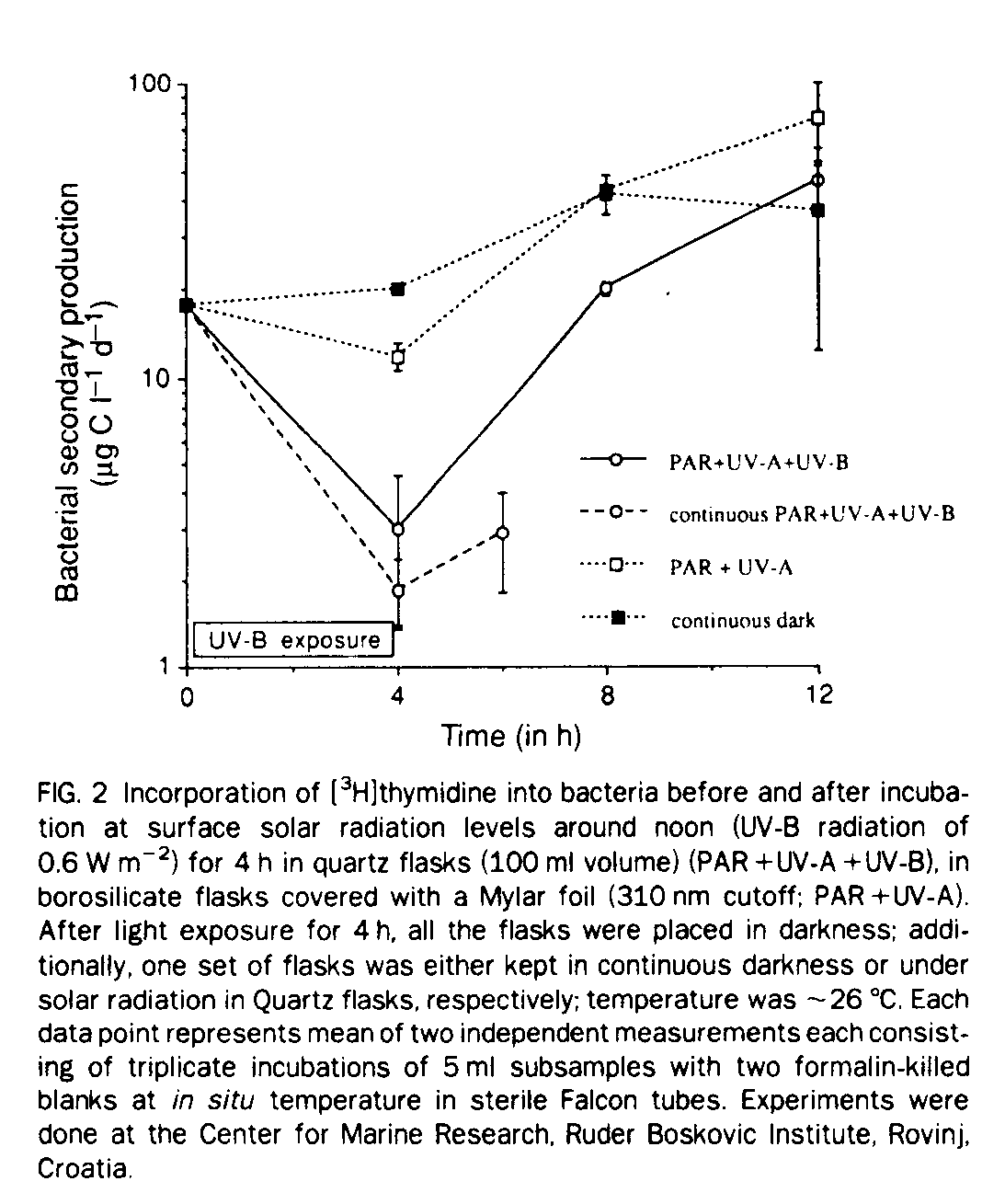
In a stratified water column mixing is restricted and bacterioplankton in the surface waters may be exposed to high levels of UV-B during their entire life cycle, whereas bacterioplankton from the subpycnocline waters never experience noticeable levels of ultraviolet radiation. We addressed this problem by exposing surface water from a highly stratified water column to surface solar radiation levels on cloudless days for 4 h. Before and immediately after exposure to natural radiation levels, thymidine incorporation into bacterial DNA was measured as well as 4 and 8 h after exposure to solar radiation (Fig. 2). After 4 h of exposure to surface light regimes, about 48% of suppression in bacterial growth is caused by UV-B and ~40% of the observed suppression is caused by photosynthetic active radiation (PAR) plus UV-A. After 4 h in dark, bacterial production in the bottle kept continuously in the dark and in the quartz bottle covered by a Mylar foil (PAR+UV-A) reached almost identical rates, indicating rapid recovery from light exposure in the absence of UV-B (Fig. 2). In the incubation flasks exposed additionally to UV-B radiation, thymidine incorporation was suppressed by about 56% even after 4 h in darkness, indicating prolonged suppression of bacterial growth after UV-B exposure.
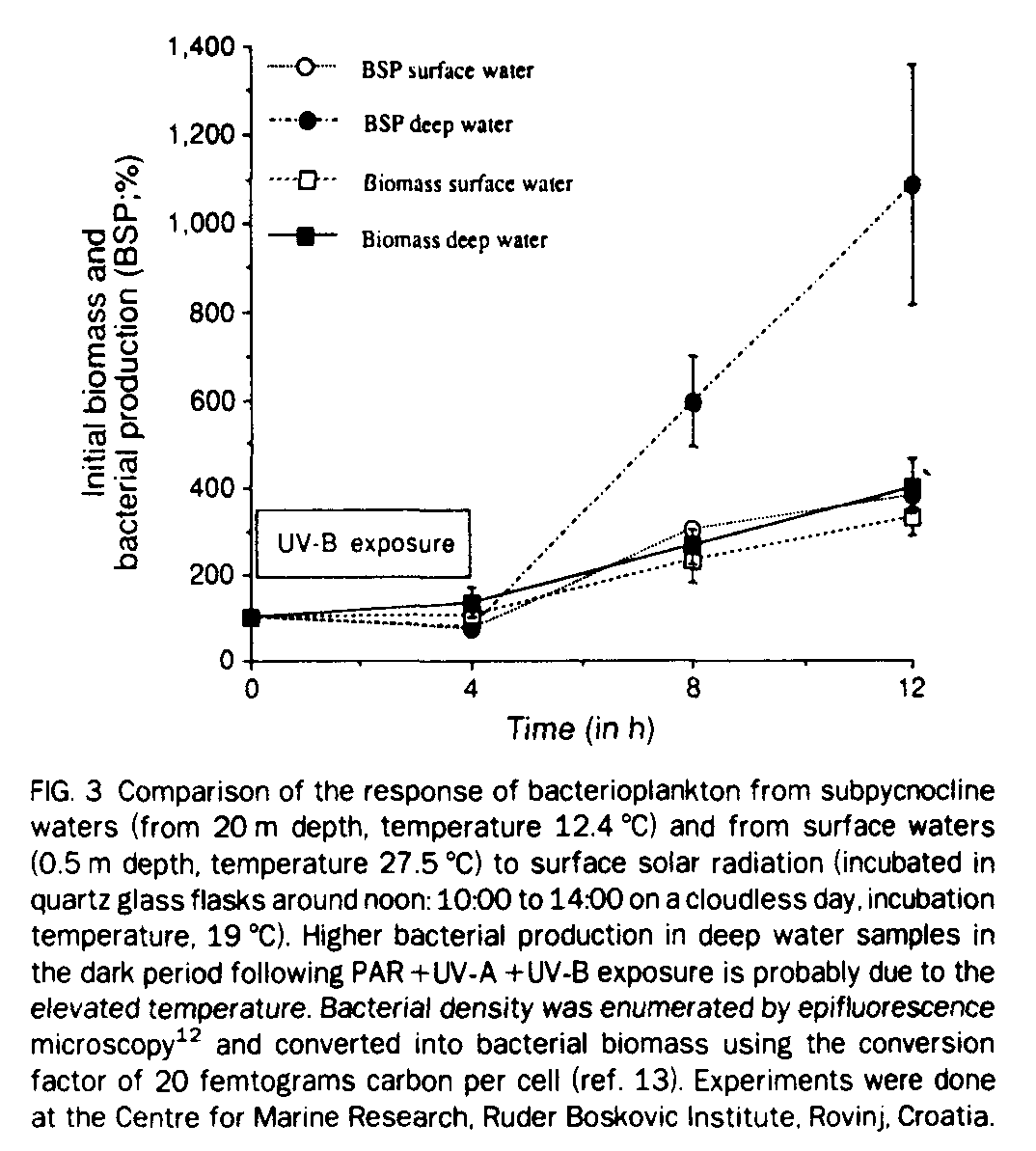
To concentrate on possible adaptations to high UV-B radiation levels, we incubated water from the subpycnocline layer (20 m depth) and compared the rate of thymidine incorporation into bacterial cells under surface water light regimes (PAR+UV-A+UV-B) with thymidine incorporation rates of surface layer bacteria (Fig. 3). After exposure for 4 h, no significant difference was detectable in thymidine incorporation rates between subpycnocline waters and surface waters; we therefore conclude that surface water bacterioplankton does not exhibit any adaptations to UV-B radiation despite the fact that they are exposed daily to a certain level of UV-B.
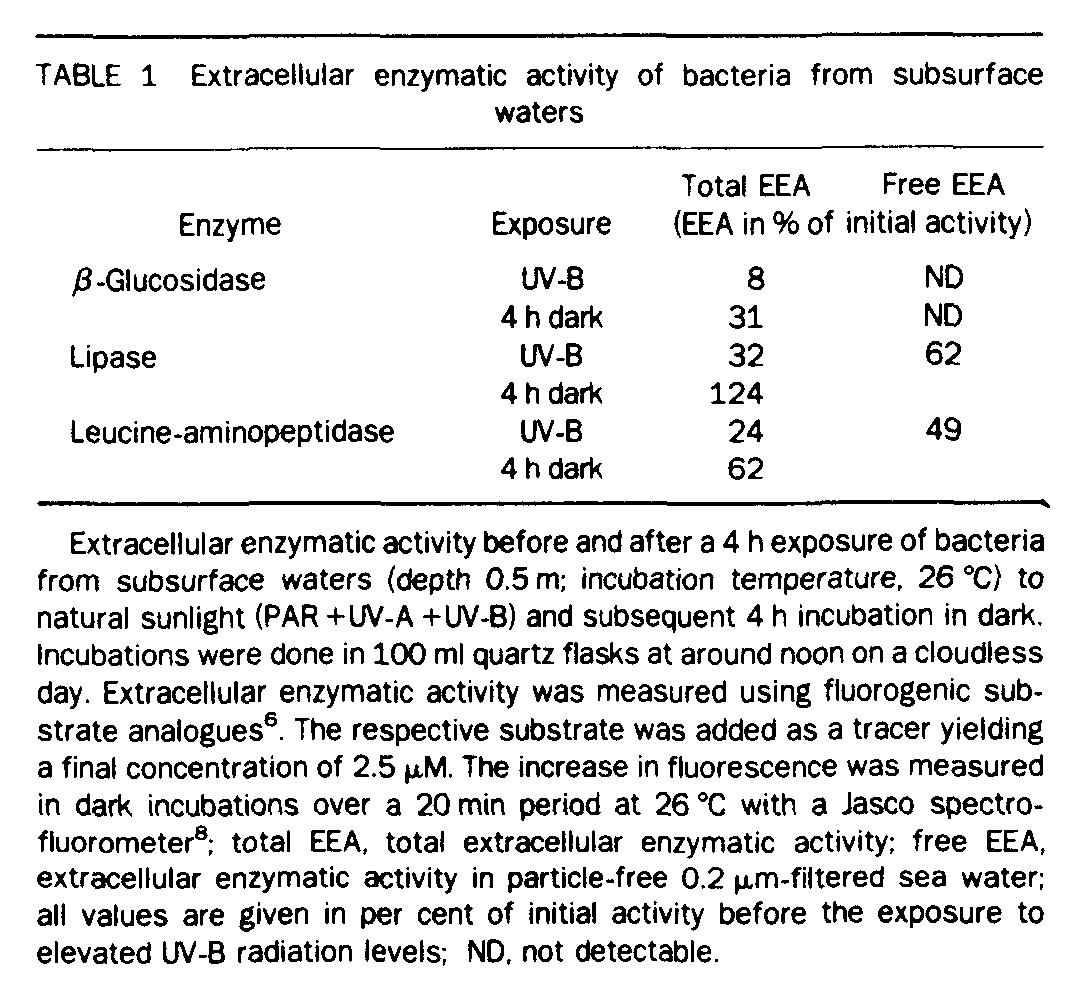
Additionally, bacterial extracellular enzymatic activity was measured using fluorogenic substrate analougus [6]. After exposure to surface solar radiation levels for 4 h, bacterial extracellular enzymatic activity was reduced to less than one third of the value measured before exposure, whereas dissolved enzymatic activity in particle-free water was reduced to 50-60% of the values before UV-B exposure, depending on the enzymes tested (Table 1). These results indicate that enzyme expression is only repressed to a minor extent; more importantly, the enzymes are photolytically cleaved during UV-B exposure[4]. Therefore UV-B radiation suppresses bacterial activity and concurrently photolytically cleaves macromolecules and thereby substitutes bacterial enzymatic activity in cleaving dissolved organic matter (DOM).
Bacterial metabolism is closely coupled to primary production[5]. Consequently, bacterial activity is highest in the upper mixed layer of the ocean where UV-B also penetrates at radiation levels high enough to influence bacterial growth. Extrapolating our measurements on UV-B penetration, we calculate that UV-B influences bacterial activity down to a depth of 10 m at around noon. Furthermore, the 1% UV-B radiation level in tropical waters is at about 30 m depth at around noon. An inhibition of bacterial activity by about 30 to 40% (depending on whether thymidine or leucine incorporation techniques have been used) is noticeable at a UV-B radiation of 0.4 W m-2; this radiation has been measured between 10:00 and 14:00 off Belize on a cloudless day down to a depth of 3.5 m. This finding has several implications in evaluating the role of bacteria in the sea. Because even surface water bacterioplankton communities from highly stratified water columns do not have adaptative capacities to protect the cells from harmful UV-B (as shown in Fig. 3), suppression of bacterial growth due to UV-B penetration in the surface layer of the ocean should be considerable. The lack of pigmentation in bacterioplankton is surprising as phytoplankton species are able to synthesize pigments to protect the cells from harmful radiation[1,7].
The increase in global UV-B radiation due to ozone depletion in the stratosphere might lead not only to enhanced photolysis of macromolecules in surface waters[4] but also to a suppression of the activity of the principal consumers of this photolytically cleaved DOM, and ultimately to an increase in potentially utilizable DOM in the surface layer of the ocean. The pathway of this DOM modified by increased UV-B radiation, however, remains speculative.
1. Worrest, R. C. & Hader, D. Environ. Conserv. 16, 261-263 (1989).
2. Karentz D. & Lutze, L. H. Limnol. Oceanogr. 35, 549-561 (1990).
3. Kieber, D. J., McDaniel, J. & Mopper, K. Nature 341, 637-639 (1989).
4. Mopper, K. et al. Nature 353, 60-62 (1991).
6. Hoppe, H.-G. Mar. Ecol. Prog. Ser. 11, 299-308 (1983).
7. Marchant, H. J., Davidson, A. T. & Kelly, G. J. Mar. Biol. 109, 391-395 (1991).
8. Karner, M. & Herndl, G. J. Mar. Biol. 113, 341-347 (1992).
9. Fuhrman, J. A. & Azam, F. Mar. Biol. 66, 109-120 (1982).
10. Robarts, R. D. & Wichs, R. J. Limnol. Oceanogr. 34, 213-222 (1989).
11. Simon, M. & Azam, F. Mar. Ecol. Prog. Ser. 51, 201-213 (1989).
12. Hobbie, J. E., Daley, R. J. & Jasper, S. Appl. Environ. Microbiol. 33, 1225-1228 (1977).
13. Lee, S. & Fuhrman, J. A. Appl. Environ. Microbiol. 53, 1298-1303 (1987).
{kind=link}
{kind=link}
{kind=link}
{kind=link}