 Reproduced, with permission, from:
Teramura, A. H. and J. H. Sullivan. 1991. Potential impacts of increased solar UV-B on global plant productivity. In Photobiology, ed. E. Riklis, 625-34. New York: Plenum Press.
Reproduced, with permission, from:
Teramura, A. H. and J. H. Sullivan. 1991. Potential impacts of increased solar UV-B on global plant productivity. In Photobiology, ed. E. Riklis, 625-34. New York: Plenum Press.
Department of Botany, University of Maryland
College Park, Maryland 20742 U.S.A.
Most of our knowledge of the effects of UV-B radiation on plants comes from studies of economically important crops. One species which has been extensively studied is soybean. Two cultivars grown for six seasons under enhanced UV-B radiation offered contrasting sensitivities with Essex exhibiting reductions in yield of 19 to 25% in four of the years, while Williams, was unaffected by increased UV-B radiation. The effectiveness of UV-B radiation was strongly affected by prevailing microclimatic factors such as precipitation patterns and air temperature.
In one of very few studies which examined UV-B radiation sensitivity in forest species, one important conifer species, loblolly pine, has been shown to be particularly sensitive in greenhouse tests. Ongoing field studies suggest that they also respond to UV-B radiation even under a full solar spectrum by increasing needle flavonoid content. However, the long-term effects have not yet been determined.
Plants have developed natural adaptations such as anatomical, morphological and biochemical changes which protect them from UV-B radiation. The extent of these natural adaptations may be related to the geographic origin of the species. It has been hypothesized that species originating from areas which receive high levels of UV-B radiation would be highly resistant to UV-B radiation. Plants collected along a 3000 m elevational gradient in Hawaii; showed differences in sensitivity which were correlated with elevation. Most plants native to low elevations were sensitive to UV-B, but plants from the higher elevations, where UV-B is greatest, were very tolerant to UV-B radiation.
Scientists have now accumulated sufficient evidence to show that human activity is rapidly changing the chemical composition of trace gases in the earth's atmosphere. Although subtle, these changes are beginning to produce global effects on the earth's ozone layer, climate and tropospheric chemistry. Of primary concern are the gases carbon dioxide (CO2), chlorofluorocarbons (CFCs), methane (CH4) and nitrous oxide (N2O) (WMO, 1986). Annual rates of increase in concentrations have been measured at 0.5% for CO2, between 5 and 7% for CFCs, 1% for CH4, 0.2% for N2O (WMO, 1986).
One important consequence of this rapid increase in trace gases results from the fact that they are all greenhouse gases. That is, they lead to an increase in global mean temperatures by absorbing infrared radiation. In addition to their contribution to global warming, chlorofluorocarbons (CFCs) may deplete the earth's protective stratospheric ozone layer. Their long atmospheric lifetimes (up to 150 years) allow them to be transported to the stratosphere where they are photodissociated and release chlorine, which catalytically destroys ozone.
Following an 18-month review, the International Ozone Trends Panel recently (March 1988) released its conclusive findings that significant global ozone depletion has already occurred. The panel cited a depletion of between 1.7% and 3.0% from 1969 to 1986 at latitudes between 30 and 64deg. North (where measurements are most extensive). The panel found that this decrease is conclusively linked to atmospheric chlorine, and is in addition to the natural variation in ozone levels (NASA, 1988).
At this time it is impossible to project future losses of ozone which will be associated with the inevitable increases in stratospheric chlorine and biogenically produced gases. Therefore, despite good prognosis for the cessation of ozone depletion based on the Montreal Protocol, which will limit CFC use, it would be premature to consider the threat of depletion to be ended.
Depletion of the ozone layer is of concern because the stratospheric ozone column is the primary attenuator of solar ultraviolet-B radiation (UV-B region, between 290 and 320 nm). A decrease in this ozone column would lead to increases in UV-B reaching the earth's surface. Though representing only a small fraction of the total solar electromagnetic spectrum, UV-B has a disproportionately large photobiological effect. One reason is that UV is readily absorbed by important macromolecules such as proteins and nucleic acids (Giese, 1964). Therefore, it is not surprising that both plant and animal life are greatly affected by increases in UV-B radiation penetrating to the earth's surface.
There exists tremendous variability in plant species sensitivity to UV-B radiation (Tevini et al., 1981; Teramura, 1983). Some species show sensitivity to present levels of UV-B radiation (Bogenrieder and Klein, 1978) while others are apparently unaffected by rather massive UV enhancements (Becwar et al., 1982). This issue is complicated further by reports of equally large response differences among cultivars of a species (Biggs et al., 1981; Teramura and Murali 1986). Approximately two-thirds of some 300 species and cultivars tested appear to be susceptible to damage from increased UV-B radiation.
Another important feature, the quality of crop yield, has only been quantitatively examined in a few species including tomato, potato (Biggs and Kossuth, 1978) and sugar beet (Ambler et al., 1978). The effectiveness of UV-B irradiation on plant growth varies seasonally and is affected by microclimate and soil fertility. For example, under water stress (Murali and Teramura, 1986) or mineral deficiency (Murali and Teramura, 1985), soybeans are less susceptible to UV-B radiation but under low levels of visible radiation sensitivity increases (Mirecki and Teramura, 1984; Warner and Caldwell, 1983). Thus field validation studies conducted over several growing seasons are crucial in any UV-B impact assessment of agricultural productivity.
In 1981, two soybean (Glycine max (L) Merr.) cultivars were chosen for study based upon preliminary greenhouse trials for UV sensitivity and planted into the field (Teramura and Murali, 1986). Based upon overall growth performance in the greenhouse, Essex was found to be sensitive while Williams was tolerant to UV-B radiation. Field experiments were conducted during May through October of 1981 to 1986 at the Agricultural Research Center, USDA, Beltsville, Maryland, U.S.A.
Supplemental UV-B radiation was supplied by filtered Westinghouse FS-40 sunlamps oriented perpendicular to the planted rows and suspended above the plants. Lamps were filtered either with 0.13 mm thick cellulose acetate (transmission down to 290 nm) for supplemental UV-B radiation or 0.13 mm Mylar Type S plastic films (absorbs all radiation below 320 nm) as a control. The radiation filtered through the cellulose acetate supplied a weighted daily supplemental irradiance of either 3.0 or 5.1 effective kJ m[-2] UV-BBE using the generalized plant response action spectrum (Caldwell, 1971) normalized to 300 nm. Plants beneath these cellulose acetate filtered lamps received supplemental doses in addition to ambient levels of UV-B radiation. These increased levels of UV-B radiation (supplemental + ambient) were similar to those which would be received at College Park, Maryland, U.S.A. (39deg. N) with anticipated 16 and 25% stratospheric ozone reductions during a cloudless day on the summer solstice (Green et al., 1980). The weighted irradiance of Mylar filtered lamps was 0, so plants beneath these lamps received only ambient levels of UV-B (8.5 effective kJ m[-2] UV-BBE on the summer solstice). Spectral irradiance beneath the lamps was measured with an Optronics Model 742 spectroradiometer equipped with a double monochromator with dual holographic grating and interfaced with a Hewlett Packard 85 printing calculator. The Spectroradiometer was calibrated using a National Bureau of Standards traceable 1000 W tugsten halogen lamp and wavelength alignment checked with known mercury emission lines using a Hg Arc lamp.
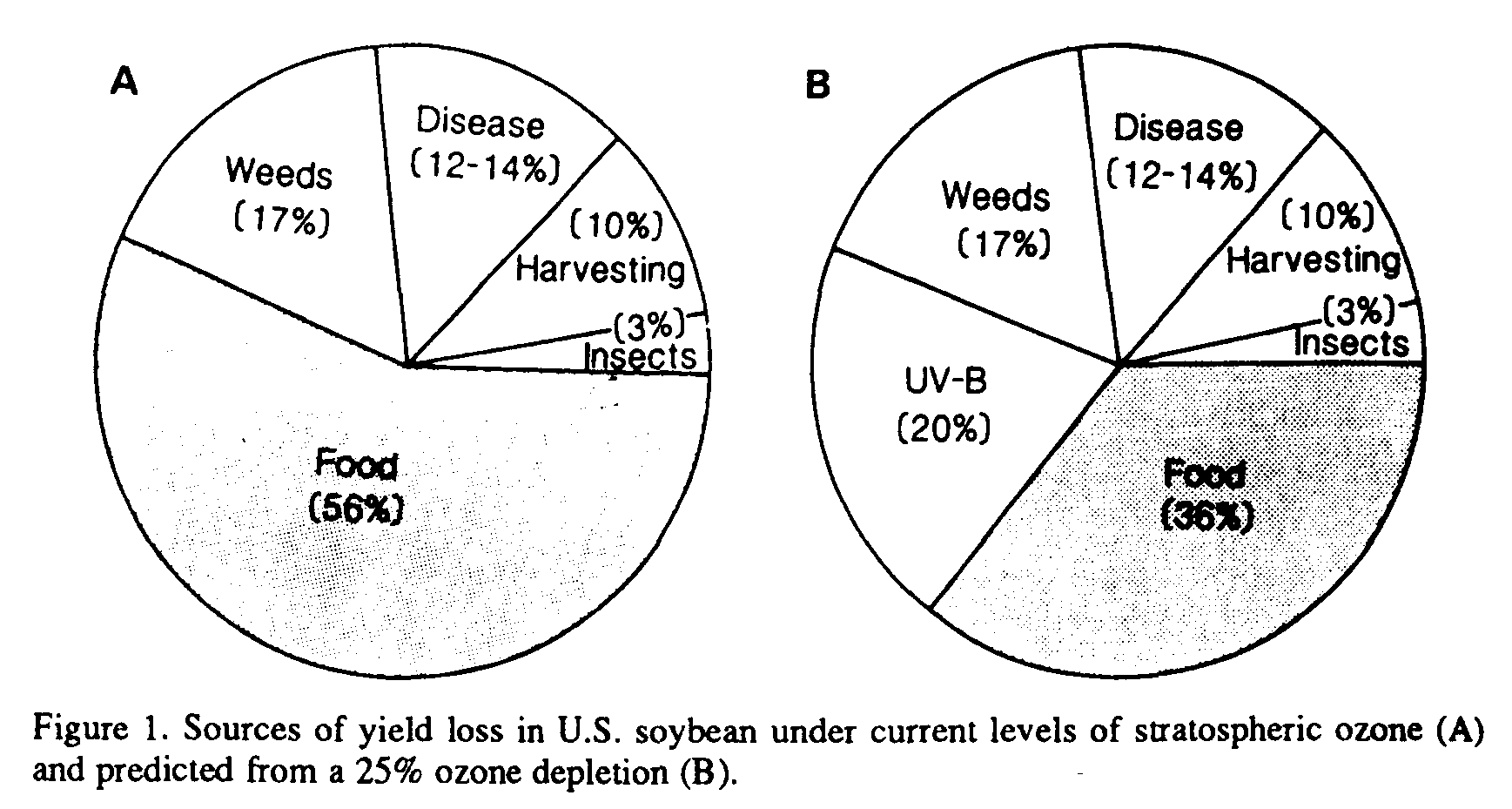
The results of this 6-year field study demonstrate intraspecific differences in UV-B sensitivity in soybean yield and quality (Table 1). However, the expression of these sensitivity differences to UV-B radiation was altered by other prevailing microclimatic factors. For the sensitive soybean cultivar Essex, a 25% ozone reduction reduced overall yield by 19-25% during 4 of the 6 years. The 1983 and 1984 seasons were characterized as hot and dry with prolonged periods of drought. Parallel field studies have shown that the effects of UV-B radiation can be masked by drought-induced growth reduction (Murali and Teramura, 1986). For perspective, Figure 1A shows the sources of yield losses in U.S. soybean under current levels of stratospheric ozone. Figure 1B shows the relative yield loss predicted from a 25% ozone depletion, indicating that UV-B-induced losses may severely limit soybean productivity. In contrast, yield increased from 5 to 22% in 5 of the 6 years for the UV-B resistant cultivar Williams.
The number of precipitation events, air temperature, the number of days of low irradiance, and UV-B radiation all interact in a complex manner to affect crop yield. Although these various interactions are complex, linear models can predict crop yield reasonably well. Based on maximum improvement of R2 (SAS 1982), the most appropriate predictive model for yield in Essex was
Y = 39.42 - 0.32A - 1.38B - 0.79C
where Y = predicted seed yield, A = total UV-B dose, B = number of days where air temperatures exceeded 35deg.C, and C = number of precipitation events. The model which best approximated yield in Williams was
Y = 154 - 0.78B - 3.08C - 13.71D
where D = precipitation frequency. Expanding models beyond three variables did not significantly increase the F-statistic or the coefficient of determination. For both cultivars R2 values exceeded 0.94 and predicted yield closely approximated actual yield. The model for Essex, the UV-susceptible cultivar, includes total UV dose while the model for the resistant Williams cultivar does not. Both models are capable of predicting crop yield within 95% confidence intervals for the five-year period (1986 was excluded from analysis because artificial irrigation was used). Through the use of such models it may be possible to more realistically assess the effects of increased levels of UV-B in concert with climatic changes than by a simple UV dose-response relationship, which does not adequately explain field observations. This interaction between UV-B dose and microclimatic variation needs further evaluation. Global climate changes altering the temperature and pattern of precipitation have been predicted due to increases in greenhouse gases in the atmosphere resulting from human activity (Titus, 1986). Such changes in climate in concert with increasing levels of UV-B radiation reaching the earth could profoundly influence the productivity of soybean and other crops, as well as that of native plant communities.
In contrast to crops, few studies have been undertaken on woody perennials (trees), which account for up to 80% of global net primary productivity and occupy as much as one-third of the land area of the United States (Whittaker, 1975; Solomon and West, 1986). Only two studies have been completed under field conditions and one of these examined the effects of solar UV-B radiation exclusion (Bogenrieder and Klein, 1982). Exclusion of naturally occurring UV-B radiation increased the growth of four broadleaf species but supplemental UV-B irradiation had no effect on growth in either Engelmann Spruce or lodgepole pine (Kaufmann, 1978). Three additional studies have demonstrated deleterious effects of UV-B radiation on tree growth and physiology (Kossuth and Biggs, 1981; Sullivan and Teramura, 1988 and 1989). However, none of these studies were extended beyond a single growing season, thus the long-term effects of UV-B radiation on trees is unknown.
The examination of UV-B radiation effects on perennial species provides a unique opportunity to observe more subtle responses to protracted UV exposure which are possible to investigate in annual species. For instance, it is presently unknown whether UV-B repair mechanisms can mitigate UV-B damage during the dormant period when ambient levels of solar UV-B are at their seasonal minimum or whether physiological changes, which accompany tissue hardening prior to entering dormancy, modify its sensitivity to subsequent UV-B exposure. The range of responses observed both inter- and intra-specifically suggests that extrapolations between annual and perennial species may not be feasible. Therefore, some direct field validation experiments on key forest species are essential before realistic estimates of the effects of increased UV-B on these species can be made.
Due to their economic importance and widespread global distribution, conifers have been selected for study in over half of the studies on trees. Some 15 species of conifers have been tested to date for susceptibility to UV-B radiation (Table 2). Of these, 7 were deleteriously affected, 5 were resistant, and 3 were favored by UV-B radiation (Sullivan and Teramura, 1988a).
Loblolly pine was one of the most susceptible species with reductions of biomass and height of 40 and 16%, respectively. Loblolly pine is the leading commercial species in the southeastern United States and accounts for a large portion of the United States pulp producing capacity (Harlow and Harbar, 1969; Walker, 1980). Therefore, there would be enormous economic consequences should increasing levels of UV-B substantially reduce loblolly pine productivity.
Kossuth and Biggs (1981) evaluated the effects of UV-B on loblolly pine growth in growth chambers and Sullivan and Teramura (1988) observed significant UV-B effects under unshaded greenhouse conditions. Both studies demonstrated that newly emergent loblolly pine seedlings were deleteriously affected by increased levels of UV-B radiation. Additionally Sullivan and Teramura (1989) have shown that established saplings are also affected in both greenhouse and field conditions by increases in UV-B radiation. In these greenhouse studies on one-year old saplings, loblolly pine responded in a dose-specific manner to UV-B radiation by increasing needle flavonoid concentrations. The rate of increase was also dose-specific, and at a low supplemental UV-B dose (11 kJ m[-2]) the increases appeared only after photosynthetic capacity and growth had been reduced. Rapid increases in flavonoid concentration initially protected plants from deleterious UV-B effects at the highest supplemental dose (19 kJ m[-2]). However, this protection was incomplete and after six months of irradiation, needle flavonoid concentrations decreased, producing reductions in photosynthetic capacity and growth.
Due to the nature of greenhouse environments, however, caution must be exercised in the extrapolation of these results to the field setting. The effectiveness of UV-B is often exaggerated under growth chamber and greenhouse conditions due to reduced photosynthetic photon flux density (PPFD between 400 and 700 nm) levels (Mirecki and Teramura, 1984; Warner and Caldwell, 1983). This apparent increase in UV-B sensitivity in controlled environments is probably due to reductions in photoprotective and photoreactivation mechanisms. Preliminary results from a field study on loblolly pines demonstrate that needle flavonoid content was increased and growth in one of four local seed sources was reduced under supplemental UV-B radiation simulating a 25% ozone reduction. These results demonstrate that even under a complete solar spectrum, loblolly pines were sensitive to UV-B radiation and responded by increasing needle flavonoid concentrations. These increases could reduce further sensitivity but the long-term protective role of flavonoids has not yet been demonstrated. Studies extending over several growing seasons and an evaluation of the metabolic or energetic costs of producing and maintaining high flavonoid concentrations will be necessary to determine the effects of increasing solar UV-B radiation on overall tree productivity.
Currently, almost no information exists on the degree of variability that exists in our natural vegetation and therefore we have little basis upon which to assess the potential impacts of increasing levels of UV-B on natural plant communities. To address the broad question of potential global UV effects on terrestrial communities and ecosystems, we must currently make the unlikely assumption that native perennial woody trees and shrubs respond in a fashion analogous to annual herbaceous crop species. An additional function to studying UV adaptations in natural plant species is that we might find novel or unique protective mechanisms which have not been detected in crop plants already exposed to intensive artificial selection. These might then be useful in future crop breeding programs after careful screening and genetic analysis.
Perhaps the first place to search for UV-B responsiveness in native plants is in regions where natural levels of UV-B are already quite high. Since the weighted daily UVB dose received at low latitude, high elevation sites (i.e. tropical mountains) can be nearly six-fold greater than the maximum dose received at arctic latitudes (Caldwell et al., 1982), plants which naturally occur in such high UV environments would undoubtedly have evolved specific adaptations which protect them from the deleterious effects of UV-B (Antonovics, 1975). For example, Caldwell et al. (1982) found that arctic ecotypes (variants of the same species) of Oxyria digyna were consistently more sensitive to UV radiation than their counterparts collected from mountains in lower latitudes. The presence of secondary compounds such as flavonoids which may act as solar screens, absorbing UV and not allowing it to reach sensitive tissue layers in the epidermis of leaves has been shown to vary greatly among various plant species collected along a latitudinal gradient (Robberecht and Caldwell, 1978). However, a simple correlation between UV sensitivity and epidermal flavonoid concentrations does not seem to exist (Barnes et al., 1987). Caldwell et al., (1982) and Barnes et al. (1987) have also reported that the differences in sensitivity between plants collected from high UV environments (tropical mountains) were not simply due to differences in epidermal flavonoids. Instead, it appeared that the photosynthetic apparatus of these high elevation tropical plants was inherently more resistant to UV than that of plants collected from higher latitudes. However, the specific nature of these inherent differences has yet to be elucidated.
By limiting our studies to crop species in temperate environments, we may be observing only a small fraction of the range of UV protective mechanisms that plants have evolved. Therefore, it would be particularly productive to examine the types or adaptations possessed by plants naturally growing in regions of the world with the greatest solar UV-B flux. In a recent study, seeds were collected from 132 native and introduced plant species growing over a 3,000 m elevational gradient in Hawaii (23deg.N latitude). Seeds were brought back to the University of Maryland, U.S.A. and germinated in the greenhouse under artificial UV lamps simulating 20 and 40% ozone depletions over Honolulu, Hawaii during clear sky conditions on the summer solstice (14.6 and 22.2 kJ m[-2] effective UV-BBE according to an empirical model of Green et al. 1950). In the UV-B radiation treatment simulating a 40% ozone depletion, only 8% of the species collected from sea level to 500 m were tolerant to UV, while tolerance increased markedly in species collected from higher elevations (Table 3). All species collected above 2,000 m were found to be tolerant of UV-B radiation levels simulating a 40% ozone depletion, implying that these plants have adapted to the high levels of UV presently incident at these sites. A greater understanding of the range and types of natural UV protective mechanisms available to plants would provide a better basis upon which to make predictions of the effects of increased levels of solar UV on natural plant communities.
Antonovics, J. (1975) Predicting evolutionary response of natural populations to increased UV radiation. In. Climatic Impact Assessment Program (CIAP), Monograph 5 (Edited by Nachtwey, D.S., M.M. Caldwell and R.H. Biggs), pp. 8-7--8-27. U.S. Dept. Transport, Report No. DOT-TST-75-55, National Techn. Infor. Serv., Springfield, Virginia.
Barnes, P.W., S.D. Flint and M.M. Caldwell (1987) Photosynthesis damage and protective pigments in plants from a latitudinal arctic/alpine gradient exposed to supplemental UV-B radiation in the field. Arctic and Alpine Research 19,21-27.
Becwar, M.R., F.D. Moore III and M.J. Burke (1982) Effects of deletion and enhancement of ultraviolet-B (280-315 nm) radiation on plants grown at 3000 m elevation. J. Amer. Soc. Hort. Sci. 107,771-779.
Biggs, R.H., S.V. Kossuth and A.H. Teramura (1981) Response of 19 cultivars of soybeans to ultraviolet-B irradiance. Physiol. Plant. 53,19-26.
Biggs, R.H. and S.V. Kossuth (1978) Effects of ultraviolet-B radiation enhancement under field conditions on potatoes, tomatoes, corn, rice, southern peas, peanuts, squash, mustard and radish. UV-B Biological and Climatic Effects Research (BACER), Final Report, EPA, Washington, D.C.
Bogenrieder, A. and R. Klein (1978) Die abhangigkeit der UV-empfindlichkeit von der lichtqualitat bei der aufzucht (Lactuca sativa L.). Angew. Botanik 52,283-293.
Caldwell, M.M., R. Robberecht, R.S. Nowak and W.D. Billings (1982) Differential photosynthetic inhibition by ultraviolet radiation in species from the arctic-alpine life zone. Arctic and Alpine Res. 14,195-202.
Caldwell. M.M. (1971) Solar UV irradiation and the growth and development of higher plants. In Photophysiol. VI (Edited by A.C. Giese), pp. 131-177.
Giese, A.C. (1964) Studies on ultraviolet radiation action upon animal cells. In Photophysiology Vol. 2, (Edited by A.C. Giese), pp. 203-245. Academic Press, NY-London.
Green, A.E.S., K.R. Cross and L.A. Smith (1980) Improved analytical characterization of Ultraviolet skylight. Photochem. Photobiol. 31,59-65.
Harlow, W.M and E.S. Harbar (1969) Textbook of Dendrology. pp. 87-90, McGraw-Hill, ISBN 07-026569-0.
Kaufmann, M.R. (1978) The effect of ultraviolet (UV-B) radiation on Engelmann spruce and lodgepole pine seedlings. UV-B Biological and Climatic Effects Research (BACER). Final Report EPA-IAG-D6-0168. EPA, Washington, DC.
Kossuth, S.V. and R.H. Biggs (1981) Ultraviolet-B radiation effects on early seedling growth of Pinaceae species. Can. J. For. Res. 11,243-248.
Mirecki, R.M. and A.H. Teramura (1984) Effects of ultraviolet-B irradiance on soybean. The dependence of plant sensitivity on the photosynthetic photon flux density during and after leaf expansion. Plant Physiol. 74,475-480.
Murali, N.S. and A.H. Teramura (1986a) Effectiveness of UV-B radiation on the growth and physiology of field-grown soybean modified by water stress. Photochem. Photobiol. 44,215-220.
Murali, N.S. and A.H. Teramura (1985) Effects of ultraviolet-B irradiance on soybean. VI. Influence of phosphorus nutrition on growth and flavonoid content. Physiol. Plant. 63,413-416.
National Aeronautics and Space Administration (NASA) (1988) Executive Summary of thc Ozone Trends Panel, NASA, Washington, D.C.
Robberecht, R. and M.M. Caldwell (1978) Leaf epidermal transmittance of ultraviole radiation and its implication for plant sensitivity to ultraviolet-radiation induced injury. Oecologia 32,277-287.
Solomon, A.M. and D.C. West (1986) Simulating forest responses to expected climate change in eastern North America: Applications to decision-making in the forest industry. In Climate Change and Future Forest Management in the United States (Edited by W.E. Shands). The Conservation Foundation, Washington, D.C. In press.
Sullivan, J.H. and A.H. Teramura (1988) Effects of ultraviolet irradiation on seedling growth in the pinaceae. Amer. J. Bot. 75,225-230.
Sullivan, J.H. and A.H. Teramura (1989) The effects of ultraviolet-B radiation on loblolly pine: 1. Growth, photosynthesis and pigment production in greenhouse-grown saplings. Physiol. Plant 77, 202-207.
Teramura, A.H. and N.S. Murali (1986) Intraspecific differences in growth and yield of soybean exposed to ultraviolet-B radiation under greenhouse and field conditions. Environmental and Experimental Botany 26,89-95.
Terarnura, A.H. (1983) Effects of ultraviolet-B radiation on the growth and yield of crop plants. Physiol. Plant. 58, 415-422.
Tevini, M., W. Iwanzik and U. Thoma (1981) Some effects of enhanced UV-B irradiation on the growth and composition of plants. Planta 153,388-394.
Titus, J.G. (1986) The causes and effects of sea level rises. In Effects of Changes in Stratospheric Ozone and Global Climate. VI. (Edited by J.G. Titus) pp. 219-248. U.S. Environmental Protection Agency. Washington, D.C.
Walker, L.C. (1980) The southern pine region. In Regional Silviculture of the United States. (Edited by J.W. Barrett). John Wiley and Sons, ISBN 0471-05645-6.
Warner, C.W. and M.M. Caldwell (1983) Influence of photon flux density in the 400-700 nm waveband of inhibition of photosynthesis by UV-B (280-320 nm) irradiation in soybean leaves: separation of indirect and immediate effects. Photochem. Photobiol. 38,341-346
Whittaker, R.H. (1975) Communities and Ecosystems. MacMillan Co., New York.
World Meteorological Organization (WMO) (1986) Atmospheric Ozone 1985. Assessment of our Understanding of the Processes Controlling its Present Distribution and Change, WMO Global Ozone Research and Monitoring Project, Report No. 16, WMO, Geneva, Switzerland.
{kind=link}
{kind=link}
{kind=link}
{kind=link}